
Cytopathic Effect (CPE) Evaluation in Veterinary Virology
Overview and Principles of Cytopathic Effect (CPE) Evaluation in Veterinary Virology
The systematic observation and interpretation of virus-induced morphological alterations in cultured cells--collectively termed the cytopathic effect (CPE)--constitute one of the oldest, most fundamental, and conceptually richest methodologies in veterinary virology. As a clinical pathologist approaching this subject, I must emphasize that CPE is not merely a convenient laboratory endpoint; it is a dynamic, quantitative, and mechanistically informative readout of virus-host cell interaction at the molecular, organellar, and supracellular levels. The evaluation of CPE bridges classical virology with modern cell biology, providing insights into viral pathogenesis, tropism, replication kinetics, and the efficacy of antiviral interventions. This section establishes the foundational principles governing CPE evaluation, drawing on decades of empirical observation and recent technological advances to frame a comprehensive conceptual infrastructure for the chapters that follow.
The Biological Basis of CPE: A Mechanistic Overview
At its core, CPE represents the phenotypic consequence of viral subversion of host cellular machinery. Every virus, by virtue of its obligate intracellular replicative strategy, must commandeer ribosomes, metabolic substrates, membrane compartments, and cytoskeletal elements to propagate its genome and assemble progeny virions. The visible manifestations of this hijacking--whether cell rounding, detachment, syncytium formation, cytoplasmic vacuolation, or nuclear pyknosis--reflect the specific biochemical insults inflicted by viral proteins on host structures [1, 2]. For instance, the rounding and rapid detachment observed in picornavirus-infected cells, such as those infected with Porcine Teschovirus, result from the proteolytic cleavage of host translation initiation factors and the disruption of intermediate filaments and microtubules [1]. Similarly, the fusogenic glycoproteins of paramyxoviruses, including Newcastle Disease Virus and Avian Metapneumovirus, mediate pH-independent membrane fusion between adjacent cells, producing the multinucleated giant cells (syncytia) that are pathognomonic for these infections [3, 2]. The molecular diversity of these effector mechanisms is vast, yet the fundamental principle remains constant: CPE is the integrated output of a cascade of molecular events initiated by viral entry and culminating in cellular dysfunction or death.
Critically, the induction of CPE is not a universal property of all viruses. Some notable veterinary pathogens, including certain biotypes of Bovine Viral Diarrhea Virus and the slow-growing gammaherpesviruses such as Equine Herpesvirus 5, establish persistent infections in cell culture with minimal or no morphological change [4, 5]. The distinction between cytopathic (CPE-positive) and non-cytopathic (CPE-negative) viral biotypes is of profound clinical and diagnostic significance. For BVDV, this dichotomy correlates with the presence or absence of the NS5A-NS5B cleavage product, which modulates interferon induction and apoptosis [4]. Non-cytopathic strains often establish lifelong persistent infections in cattle, escaping immune detection and serving as reservoirs for horizontal transmission. Thus, the absence of CPE in a diagnostic setting does not imply the absence of infection; it may instead indicate the need for alternative detection methods, such as immunostaining or molecular amplification, to uncover the presence of a stealth pathogen.
Viral and Cellular Determinants of CPE Morphology
The specific pattern of CPE observed in a given virus-cell system is dictated by a complex interplay between viral genetic determinants and the intrinsic properties of the host cell. Different cell lines derived from the same species can exhibit dramatically divergent susceptibilities to CPE induction by the same virus. A seminal study of Severe Fever with Thrombocytopenia Syndrome Virus infection across a panel of mammalian cell lines demonstrated that while tiger-derived TLT cells, feline CRFK and FCWF-4 cells, and porcine CPK cells all developed conspicuous CPE, Vero E6 (monkey), A549 (human), BHK-21 (hamster), and several deer- and hedgehog-derived cell lines remained morphologically intact despite supporting varying levels of viral replication [6]. This cell-type-dependent susceptibility has critical implications for virus isolation protocols: the selection of an appropriate cell line is paramount to maximize the sensitivity of CPE-based detection. Similarly, the adaptation of Newcastle Disease Virus to chicken embryo fibroblast (CEF) versus Vero cells requires differential passage numbers to achieve reproducible CPE, reflecting the molecular barriers to cross-species adaptation [3].
The three-dimensional architecture of the cytoskeleton and the integrity of intercellular junctions also modulate CPE expression. For enteric viruses such as Porcine Deltacoronavirus, infection of polarized intestinal epithelial cells (IPEC-J2) produces CPE that is both delayed and morphologically distinct from that observed in conventional kidney cell lines, likely due to the restrictive nature of tight junctions and the polarized distribution of viral receptors [7]. This observation underscores the importance of using physiologically relevant cell culture models when evaluating CPE for viruses that target specialized epithelial barriers in vivo. The metabolic state of the cell--influenced by serum concentration, confluency, and passage number--can profoundly affect the kinetics and severity of CPE. Stressed or senescent cells are often more permissive to viral replication and exhibit accelerated CPE, confounding the interpretation of antiviral assays if not rigorously controlled.
Quantitative Approaches to CPE Evaluation
Traditional CPE evaluation relied on subjective, semi-quantitative scoring by experienced microscopists, typically on a 0 to 4+ scale reflecting the percentage of monolayer involvement. While this approach remains valuable in diagnostic virology for its speed and low cost, it is inherently limited by inter-observer variability and the inability to capture subtle kinetic differences. Modern veterinary virology has increasingly adopted objective, high-throughput, and real-time methods for CPE quantification. The real-time cell analysis (RTCA) system, which measures changes in cell-substrate impedance as a surrogate for monolayer integrity, provides continuous, label-free monitoring of CPE progression [8]. In studies of West Nile Virus in Birds and St. Louis Encephalitis Virus, the RTCA-determined time to 50% impedance reduction (CIT50) was inversely proportional to the input virus dose and could distinguish between viruses with different replication rates--West Nile virus induced CPE approximately three times faster than St. Louis encephalitis virus in the same cell line [8]. This approach enables precise quantification of neutralizing antibody titers by measuring the delay in CPE onset, offering a fully automated alternative to the traditional plaque reduction neutralization test [8, 9].
Metabolic assays, such as the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) reduction assay and the resazurin-based conversion to fluorescent resorufin, provide another robust quantitative endpoint for CPE [10, 11]. These assays measure the activity of mitochondrial dehydrogenases in viable cells, allowing the calculation of percentage protection against virus-induced cell death. The resazurin method has been successfully employed to screen for inhibitors of Feline Coronavirus and FIP, identifying chloroquine, mefloquine, and hexamethylene amiloride as lead compounds with selectivity indices of 217, 24, and 20, respectively [11]. These colorimetric and fluorometric assays must be carefully validated to distinguish true CPE inhibition from compound cytotoxicity, a challenge that is elegantly addressed by integrated analytical frameworks that combine CPE observation with lactate dehydrogenase (LDH) release, MTT reduction, and antigen detection [10]. The application of atomic force microscopy (AFM) to measure cell surface roughness represents a cutting-edge, label-free alternative for detecting early CPE, particularly relevant for viruses like rabies that produce subtle morphological changes that are difficult to score by conventional light microscopy [12].
CPE in Antiviral Drug Discovery and Neutralization Testing
The CPE inhibition assay remains the cornerstone of phenotypic antiviral screening in veterinary medicine, as it captures the net effect of a compound on the entire viral replication cycle, from entry to egress, without requiring prior knowledge of the molecular target. High-throughput screening platforms, often in 96- or 384-well formats, use automated imaging or metabolic readouts to evaluate thousands of compounds for their ability to protect cell monolayers from virus-induced destruction [13]. This approach successfully identified nitazoxanide, closantel sodium, and closantel as inhibitors of Canine Parvovirus in F81 cells, with subsequent mechanistic studies revealing that these compounds act within the first two hours post-infection, likely by blocking early entry or uncoating events [13]. Similarly, the oxazole-4-carboxamide KB-2777 was identified through CPE protection assays against human coronaviruses and subsequently shown to attenuate pro-inflammatory cytokine responses, including IL-6, IFNB1, and ISG15, in infected MRC-5 cells [14].
The virus neutralization test (VNT), which measures the ability of antibody-containing sera to prevent CPE in cell culture, remains the gold standard for assessing functional immunity against numerous veterinary pathogens, including Rift Valley Fever Virus, Rabies Lyssavirus, and Avian Influenza Virus [3, 15, 12]. The endpoint of VNT is typically defined as the highest serum dilution that completely inhibits CPE in at least 50% of replicate wells (the 50% neutralizing dose, ND50). This assay is indispensable for vaccine efficacy studies and serosurveillance, as it measures antibodies capable of blocking viral entry or replication, as opposed to binding antibodies detected by ELISA that may lack neutralizing capacity [15]. A recent study comparing competitive ELISA and VNT for Rift Valley fever virus in vaccinated sheep demonstrated that while both assays correlated well at high antibody concentrations, the VNT remained sensitive at serum dilutions of 1:64 and beyond, where the ELISA had become negative, underscoring the superior functional resolution of CPE-based neutralization assays [15].
Distinguishing CPE from Cytotoxicity: The Methodological Imperative
One of the most critical and frequently overlooked challenges in CPE-based assays is the accurate differentiation between virus-induced CPE and compound-induced cytotoxicity [10]. This distinction is particularly problematic in disinfectant efficacy testing, where chemical residues may directly damage cell monolayers, producing morphological changes that mimic viral CPE and leading to false-positive assessments of antiviral activity. A rigorous multi-step framework for addressing this issue has been developed using African Swine Fever Virus as a model, integrating parallel CPE observation, LDH release assays, MTT reduction, and antigen-specific detection using lateral flow immunoassays (for p30) and immunofluorescence (for p54) [10]. The study revealed that at certain dilutions, the disinfectant-treated virus groups showed lower cytotoxicity than the disinfectant-only controls, likely due to the formation of virus-disinfectant complexes that reduced the bioavailability of cytotoxic components. This phenomenon highlights the complexity of virus-chemical interactions and the need for multiple orthogonal readouts to avoid misinterpretation. The same principle applies to natural product screening: plant extracts such as those from Zanthoxylum bungeanum (Sichuan pepper) and Dryopteris crassirhizoma must be evaluated for inherent cytotoxicity before CPE protection can be attributed to direct antiviral activity [16, 17].
CPE as a Kinetic and Dynamic Readout
The temporal evolution of CPE provides a rich source of kinetic information that is often underutilized in routine diagnostic practice. The rate of CPE development, the lag phase before its onset, and the pattern of progression across the monolayer can distinguish between viral strains with different pathogenic potentials. For example, clinical isolates of chikungunya virus from India were categorized as "fast-growing" (CPE observed within 24 hours post-infection) or "slow-growing" based on their replication kinetics in Vero cells, with fast-growing isolates associated with elevated serum levels of IFN-alpha, IL-1RA, IL-17F, IL-9, MCP-1, and MIP-1alpha in the corresponding patients [18]. This correlation between in vitro CPE kinetics and in vivo cytokine profiles suggests that CPE evaluation can serve as a surrogate marker for viral pathogenicity. Similarly, the time-of-addition assay, which involves adding a test compound at various intervals post-infection and monitoring subsequent CPE, can pinpoint the stage of the viral life cycle targeted by an antiviral agent [16, 17]. For the Zanthoxylum bungeanum extract, CPE inhibition was most pronounced when the extract was present during the early phase of HCoV-OC43 infection, leading to the identification of viral entry as the primary target, mediated by hydroxy-alpha-sanshool and p-coumaric acid [16].
Integration with Molecular Diagnostics
While CPE evaluation remains a frontline method for virus isolation and phenotypic characterization, its role in the modern diagnostic laboratory is increasingly integrated with molecular techniques. The isolation of Equine Herpesvirus 1, Equine Herpesvirus 2, and Equine Herpesvirus 5 from equine tissues in Serbia exemplifies this synergy: CPE was observed on RK-13 cells after 2-5 days for EHV-1 and EHV-5, but the final identification relied on multiplex nested PCR and sequencing of the glycoprotein B gene [5]. CPE isolation followed by molecular confirmation provides a far more definitive diagnosis than PCR alone, as it confirms the presence of replication-competent virus and allows for subsequent characterization of antigenic and genetic variants. For novel or emerging pathogens, such as the recombinant Porcine Teschovirus strain SD2023, the observation of typical CPE in PK-15 cells was the essential first step that led to full-genome sequencing and the discovery of a unique recombination event involving parental strains of PTV-4 and PTV-5 [1].
In the chapters that follow, the specific CPE patterns associated with major viral families affecting veterinary species will be explored in detail, along with the standardized protocols for CPE observation, recording, and interpretation in both diagnostic and research settings. The principles established here--mechanistic diversity, cellular determinants, quantitative rigor, and integration with molecular methods--form the conceptual foundation for all subsequent discussions of CPE in veterinary virology.
Methodological Protocols for CPE Induction and Observation in Cell Culture Systems
The induction and observation of the cytopathic effect (CPE) in cell culture systems remain the cornerstone of classical virology, providing the first tangible evidence of viral replication and cytopathogenicity. For the veterinary clinical pathologist, a rigorous, standardized, and well-documented protocol is not merely a procedural formality; it is the critical foundation upon which virus isolation, quantification, and characterization are built. This section delineates the comprehensive methodological protocols for CPE induction and observation, drawing on a wealth of contemporary research to provide an exhaustive guide for the diagnostic and research laboratory.
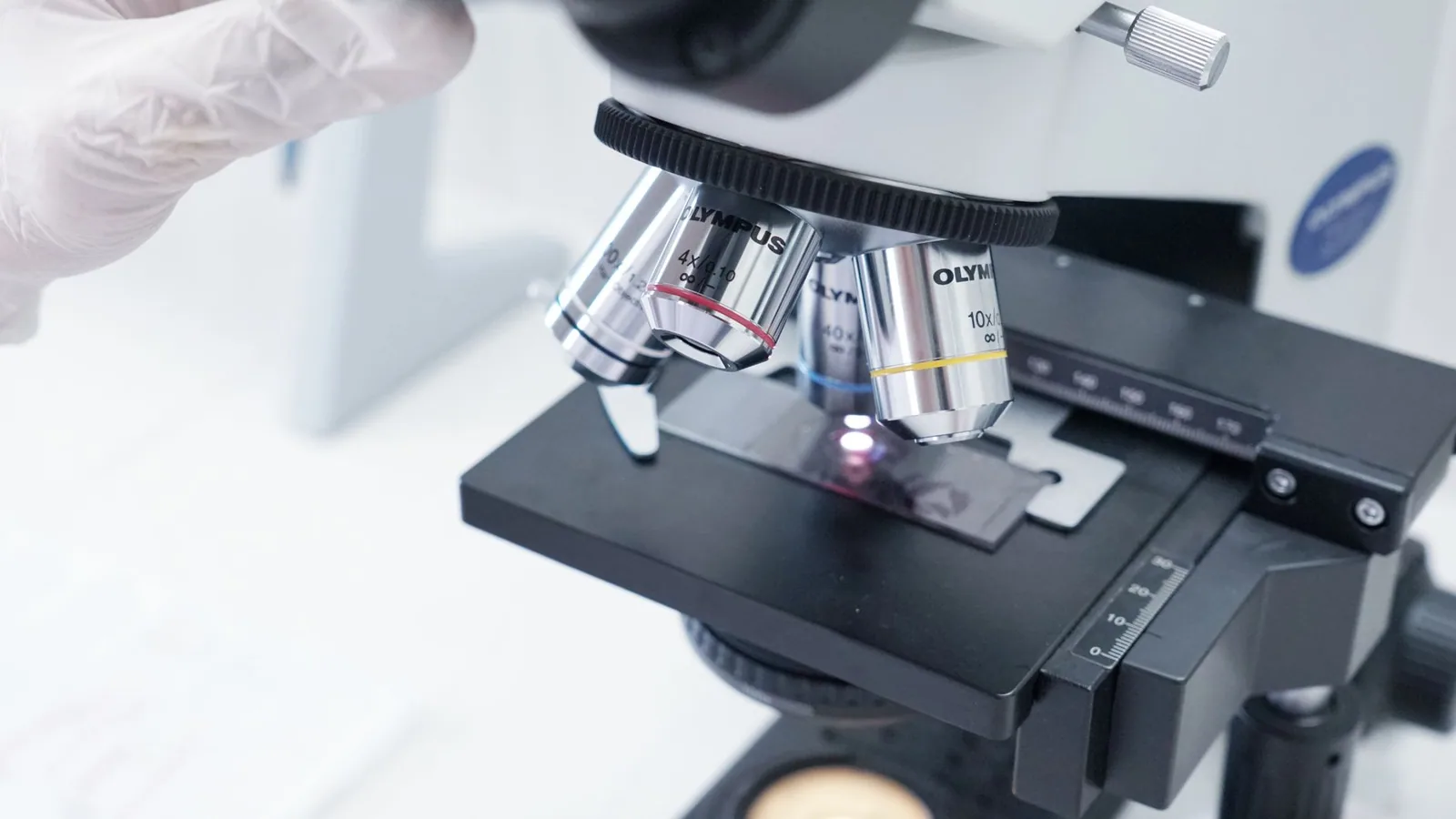
1.0 Pre-Inoculation Cell Culture Preparation and Quality Control
The success of any CPE-based assay is predicated on the health and uniformity of the cell monolayer. Prior to inoculation, cell lines must be propagated under strictly controlled conditions. The choice of cell line is paramount and is dictated by the viral pathogen of interest. For instance, the isolation of Equine Herpesvirus 1 and Equine Herpesvirus 5 is classically performed on RK-13 cells, where a characteristic CPE is observed 48-72 hours post-inoculation (hpi) [5]. Conversely, the isolation of Porcine Reproductive and Respiratory Syndrome Virus (PRRSV) requires the use of primary porcine alveolar macrophages (PAM) or the simian kidney cell line MARC-145, as these are the only cells permissive to the virus's restricted tropism [19]. Similarly, the isolation of Bovine Viral Diarrhea Virus (BVDV) is typically performed on MDBK cells, where a cytopathic (cp) biotype will induce a clear CPE, while a non-cytopathic (ncp) biotype will not [4].
Protocol for Monolayer Preparation:
- Cell Seeding: Seed cells into appropriate vessels (e.g., 24-well plates, 96-well plates, or T-25 flasks) at a density that will yield a confluent, but not over-confluent, monolayer within 24 hours. For example, for Vero E6 cells used in Severe Fever with Thrombocytopenia Syndrome Virus (SFTSV) studies, a seeding density of 2-3 x 10^5 cells/mL is standard [6].
- Medium and Incubation: Use the recommended growth medium (e.g., Eagle's Minimum Essential Medium (MEM) or Dulbecco's Modified Eagle Medium (DMEM)) supplemented with 2-10% fetal bovine serum (FBS), 100 U/mL penicillin, 100 ug/mL streptomycin, and 2.5 ug/mL amphotericin B. Incubate at 37 degrees C in a humidified atmosphere with 5% CO2.
- Quality Control (QC): Before inoculation, examine the monolayer under an inverted phase-contrast microscope. The monolayer must be >90% confluent, uniform, and free from any signs of degeneration, contamination, or spontaneous CPE. A "cell control" well, which receives only maintenance medium, must be included in every assay to serve as a baseline for normal cell morphology.
2.0 Virus Inoculation and Adsorption
The adsorption phase is a critical step where the virus is allowed to bind to and enter the host cells. The efficiency of this step directly influences the kinetics and magnitude of the subsequent CPE.
Protocol for Inoculation:
- Inoculum Preparation: Viral inocula can be derived from clinical specimens (e.g., tissue homogenates, nasal swabs, serum) or from previously titrated virus stocks. For clinical samples, it is often necessary to filter-sterilize the inoculum through a 0.22 um filter to remove bacterial and fungal contaminants [5]. The inoculum volume should be sufficient to cover the monolayer, typically 100 uL for a 96-well plate or 1 mL for a T-25 flask.
- Adsorption: Aspirate the growth medium from the cell monolayer and wash gently with phosphate-buffered saline (PBS) to remove residual serum, which can contain non-specific inhibitors. Add the viral inoculum and incubate for 1 hour at 37 degrees C with gentle rocking every 15 minutes to ensure even distribution and prevent the monolayer from drying out [5].
- Post-Adsorption: After the adsorption period, remove the inoculum and wash the monolayer once or twice with PBS to remove unadsorbed virus particles. This step is crucial for accurate quantification in plaque assays and for preventing secondary infections that could confound CPE interpretation.
- Addition of Maintenance Medium: Add fresh maintenance medium (typically MEM with 2% FBS, a lower concentration to slow cell division and enhance CPE visualization) and return the cultures to the incubator.
3.0 Systematic Observation and Grading of CPE
The observation of CPE is a dynamic process that requires meticulous, scheduled monitoring. The frequency of observation depends on the virus; for rapidly replicating viruses like Newcastle Disease Virus (NDV) or Avian Influenza Virus, CPE may be evident within 24-48 hours [3, 2]. For slower-growing viruses, such as Equine Herpesvirus 5 or some aquatic birnaviruses, observation may need to extend for 7-10 days, with blind passages performed if no CPE is initially observed [5].
Standardized CPE Grading System (0 to 4+):
- 0 (No CPE): The monolayer is intact and indistinguishable from the uninfected cell control.
- 1+ (25% CPE): Early focal areas of cell rounding, shrinkage, or granulation are visible. The monolayer is still largely intact. For example, early infection with Porcine Teschovirus in PK-15 cells presents as scattered, rounded, refractile cells [1].
- 2+ (50% CPE): The CPE is more extensive, with approximately half of the monolayer showing morphological changes. Syncytia (multinucleated giant cells) may be forming, as seen with Newcastle Disease Virus [2] or Feline Coronavirus and FIP [11]. The monolayer begins to detach from the substrate.
- 3+ (75% CPE): The majority of the monolayer is affected. Large areas of the cell sheet have detached, leaving a lace-like or "moth-eaten" appearance. This is the optimal time for harvesting virus for stock preparation or for performing a neutralization test readout.
- 4+ (100% CPE): Complete destruction of the monolayer. Virtually all cells have detached, rounded up, or lysed. The well may appear clear or contain cellular debris.
Morphological Characterization of CPE: The specific type of CPE can be highly diagnostic.
- Rounding and Detachment: This is the most common CPE, observed with picornaviruses like Porcine Teschovirus [1] and Foot-and-Mouth Disease Virus [20], as well as many flaviviruses like West Nile Virus in Birds [8].
- Syncytia Formation: Fusion of adjacent cell membranes leads to the formation of large, multinucleated giant cells. This is characteristic of paramyxoviruses (e.g., Newcastle Disease Virus [2]), some herpesviruses, and coronaviruses like Feline Coronavirus and FIP [11].
- Focal Clustering and Aggregation: Some viruses, such as Avian Influenza Virus, can cause cells to aggregate into clusters while the surrounding monolayer remains intact [2].
- Vacuolation and Foamy Appearance: This is a hallmark of certain retroviruses and reoviruses, where the cytoplasm becomes filled with vacuoles.
- Inclusion Bodies: While not visible by standard light microscopy without staining, certain viruses (e.g., Canine Distemper Virus) induce intracytoplasmic or intranuclear inclusion bodies that can be visualized with specific stains.
4.0 Confirmatory Assays and Advanced Detection
While direct observation of CPE is the primary endpoint, it must be confirmed to rule out non-specific cytotoxicity or bacterial contamination. This is especially critical when testing antiviral compounds or disinfectants, where the test agent itself may be toxic to the cells [10, 21].
4.1 Cytotoxicity Controls: In any antiviral or disinfectant assay, parallel "toxicity controls" (cells treated with the test compound alone, without virus) are mandatory. The concentration of the test agent must be below the cytotoxic threshold, defined as the concentration that causes <10% cell death compared to the untreated cell control [10, 13]. The use of colorimetric assays like MTT or LDH release can objectively quantify cell viability and differentiate true CPE from cytotoxicity [10].
4.2 Immunofluorescence (IFA) and Immunohistochemistry (IHC): IFA is a powerful tool to confirm that the observed CPE is due to the specific virus. After observing CPE, the cell monolayer can be fixed (e.g., with 80% acetone or 4% paraformaldehyde) and stained with a virus-specific monoclonal or polyclonal antibody conjugated to a fluorophore (e.g., FITC). The presence of specific viral antigens within the affected cells confirms the diagnosis. This technique is routinely used for confirming the isolation of Porcine Reproductive and Respiratory Syndrome Virus [19], Bovine Viral Diarrhea Virus [4], and Severe Fever with Thrombocytopenia Syndrome Virus [6].
4.3 Real-Time Cell Analysis (RTCA): Traditional CPE observation is subjective and endpoint-based. The RTCA system, which measures changes in electrical impedance across gold microelectrodes at the bottom of a culture plate, provides a continuous, quantitative, and objective measurement of CPE. As cells adhere and spread, impedance increases; as they round up and detach due to viral CPE, impedance decreases. This technology has been successfully applied to monitor the CPE of West Nile Virus in Birds and St. Louis encephalitis virus, providing a kinetic parameter (CIT50) that is inversely proportional to the infectious dose [8]. This method is far more sensitive than manual observation and allows for high-throughput screening of antiviral compounds.
5.0 Special Considerations for Non-Cytopathic Viruses
A critical diagnostic challenge is the presence of non-cytopathic (ncp) viral biotypes. The most prominent veterinary example is ncp Bovine Viral Diarrhea Virus, which can replicate in cell culture to high titers without causing any visible CPE [4]. To detect such viruses, an additional step is required: the "blind passage" or the use of an immunoperoxidase monolayer assay (IPMA) or IFA. In a blind passage, the cell lysate from a culture showing no CPE is passaged onto fresh cells. After 3-5 days, the cells are fixed and stained with a specific antibody. Alternatively, the culture supernatant can be tested for viral RNA by RT-PCR [4]. This principle also applies to the isolation of Equine Herpesvirus 5, which is notoriously slow to induce CPE and may require multiple blind passages before it is detectable [5].
6.0 Endpoint Determination and Harvesting
The endpoint of a CPE assay is determined by the experimental objective.
- For Virus Isolation: The culture is harvested when CPE reaches 3+ to 4+ (70-100% destruction). The flask is subjected to three freeze-thaw cycles (-80 degrees C to 37 degrees C) to lyse remaining cells and release intracellular virus. The lysate is clarified by centrifugation and stored at -80 degrees C as a virus stock.
- For Virus Quantification (TCID50): The endpoint is the dilution of virus at which 50% of the inoculated wells show CPE. This is calculated using the Reed-Muench method or the Spearman-Karber method. The assay is typically read when the CPE in the highest virus concentration wells is complete (4+), usually 5-7 days post-infection [15].
- For Neutralization Assays (VNT): The endpoint is the highest dilution of serum that completely inhibits CPE (i.e., provides 100% protection of the monolayer). The readout is typically performed when the virus control wells (virus + no serum) show 100% CPE [9, 15]. The colorimetric CPE inhibition assay, using dyes like resazurin, provides an objective, spectrophotometric readout for this purpose, eliminating the subjectivity of microscopic examination [11, 9].
Molecular Pathogenesis and Virological Mechanisms Underlying CPE Formation
The cytopathic effect (CPE) observable in permissive cell cultures represents the culmination of a complex, multi-factorial cascade of molecular events orchestrated by the invading virus. Far from being a singular, monolithic endpoint, CPE is a phenotypic readout of distinct and often concurrent pathogenic mechanisms, including the subversion of host biosynthetic machinery, the induction of programmed cell death, the manipulation of membrane integrity, and the cytoskeletal reorganization. A comprehensive understanding of these underlying virological mechanisms is paramount, as it informs not only the diagnostic interpretation of CPE morphology but also the rational design of antiviral strategies and the assessment of viral virulence. The following analysis dissects the molecular pathogenesis of CPE, drawing from a diverse array of veterinary viral pathogens to illustrate the spectrum of these mechanisms.
The Centrality of Host Macromolecular Synthesis Shutoff and Apoptotic Induction
A foundational mechanism driving CPE formation across numerous virus families is the targeted inhibition of host cell transcription and translation, coupled with the induction of apoptosis. This biphasic strategy ensures maximal viral resource allocation while simultaneously dismantling the host's antiviral defenses and facilitating viral dissemination with minimal inflammatory response.
This paradigm is strikingly illustrated by the Picornaviridae family. The isolated porcine teschovirus 5 (PTV-5) strain SD2023 induces a classic picornavirus CPE in PK-15 cells, characterized by cellular rounding, shrinkage, and eventual detachment from the monolayer [1]. The molecular underpinnings here are well-established across the enterovirus group. The viral 2A protease (2Apro) cleaves eukaryotic initiation factor 4G (eIF4G), a crucial component of the cap-binding complex eIF4F. This cleavage effectively shuts off cap-dependent host mRNA translation, while viral protein synthesis proceeds via a cap-independent, internal ribosome entry site (IRES)-mediated mechanism. Concomitantly, the viral 3C protease (3Cpro) degrades multiple host factors, including components of the transcription machinery (e.g., TATA-binding protein, RNA polymerase II), leading to a profound suppression of cellular transcription. This molecular assault, combined with the hijacking of membrane compartments for genome replication (e.g., autophagosome-like structures), triggers cellular stress responses, ultimately culminating in apoptosis. The pathogenic relevance of the SD2023 strain was confirmed in vivo, where it induced mild neurological symptoms and lymphocytic infiltration, consistent with the subclinical to mild disease often seen in field infections [1]. The highly variable regions and unique VP1 substitutions identified in SD2023 may influence receptor binding and the timing or intensity of this molecular cascade, a hypothesis that warrants further investigation.
Similarly, though acting through a distinct genetic strategy, the acquisition of a cytopathic (cp) biotype in Bovine Viral Diarrhea Virus (BVDV) is a textbook example of CPE emerging from a precise molecular lesion. Non-cytopathic (ncp) BVDV isolates establish persistent infections, while cp biotypes arise via RNA recombination or mutations that lead to the expression of a cleaved, non-structural protein NS3. In ncp infections, NS3 remains fused with its precursor NS2-3; in cp strains, the release of free NS3 is the key virulence factor. The isolation of the BVDV-1a strain HN01, which induced CPE in MDBK cells after two passages, represents the first characterization of this subgenotype in China [4]. The presence of free NS3 in cp BVDV activates a cascade of cellular events, including the induction of apoptosis via the intrinsic mitochondrial pathway, likely through the interaction with and cleavage of cellular proteins involved in survival signaling (e.g., poly(ADP-ribose) polymerase, PARP). This leads to the characteristic CPE of rounding, pyknosis, and cell death, a phenotype directly correlating with the virus's moderate virulence in experimentally infected calves, which developed fever, leukopenia, and respiratory signs [4].
Cytolytic Necrosis vs. Apoptosis: Kinetic and Morphological Determinants
While apoptosis is a common endpoint, many viruses, particularly those with highly lytic replication cycles, induce a more pronounced cytolytic necrosis. The distinction is not always absolute, with many infections exhibiting features of both programmed and necrotic cell death (necroptosis). The kinetics of CPE can be a critical clue to the dominant mechanism.
The Newcastle Disease Virus (NDV) is a prime example of a virus inducing a rapid, lytic CPE. In chicken embryo fibroblast (CEF) and DF-1 cells, NDV infection leads to marked cell fusion (syncytia formation), multinucleated giant cell formation, and eventual cellular disintegration [2]. The fusion process is driven by the viral fusion (F) protein, which, upon activation by proteolytic cleavage, mediates the merger of the viral envelope with the host cell membrane. In highly virulent strains, the F protein cleavage site contains a multibasic motif, allowing ubiquitous furin-like proteases to activate the protein, leading to systemic spread and rapid, extensive cell-to-cell fusion--a hallmark of velogenic NDV. The CPE observed in Vero and CEF cells during the adaptation of the NDV I-2 vaccine strain demonstrates that even attenuated strains retain this fusogenic capacity, albeit with slower kinetics and more localized CPE [3]. The resulting cell death is a mix of apoptosis and primary necrosis, driven by the massive membrane damage and metabolic collapse from synchronous viral replication and syncytium formation.
Conversely, other viruses exhibit a more subtle, slow-onset CPE that is predominantly apoptotic. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV), a highly pathogenic arterivirus, demonstrates this in its target cells, porcine alveolar macrophages (PAMs) and MARC-145 cells [19]. The CPE in PRRSV-infected macrophages is characterized by cellular rounding and detachment, with minimal syncytia formation. The virus's non-structural proteins, particularly nsp1beta and nsp2, are potent inhibitors of the interferon (IFN) response, allowing unchecked viral replication. The ensuing cellular stress, combined with the virus's direct modulation of pro-apoptotic (Bax, Bad) and anti-apoptotic (Bcl-2) pathways, culminates in delayed but extensive apoptosis. This mechanism is critical for viral pathogenesis, as apoptotic bodies can facilitate viral spread while subverting the inflammatory response, contributing to the persistent infections and immunosuppression characteristic of PRRSV [19].
Subversion of Autophagic Flux and Lysosomal Function
An increasingly recognized mechanism in CPE pathogenesis is the manipulation of the host autophagy-lysosome pathway. Viruses have evolved sophisticated strategies to either inhibit autophagy's antiviral role or to hijack its membrane compartments for replication.
This is particularly well-documented for coronaviruses. In the context of Porcine Deltacoronavirus (PDCoV) infection in IPEC-J2 intestinal epithelial cells, a fascinating dichotomy was observed. While the virus induced a clear CPE and viral antigen was abundant, the majority of CPE-positive cells did not exhibit TUNEL-positive (apoptotic) signals [7]. This suggests that PDCoV-induced CPE, at least in the early to mid-phase, is driven by non-apoptotic mechanisms, likely involving the disruption of cellular membrane integrity and organellar function. Coronaviruses are known to induce massive membrane rearrangements (double-membrane vesicles, DMVs) derived from the endoplasmic reticulum, which serve as the viral replication complex. This process disrupts ER homeostasis and can trigger ER stress. PDCoV, like other coronaviruses, may modulate autophagic flux. The study [7] indicates that the CPE may be more closely linked to these biophysical disruptions than to classical apoptosis. This is further supported by work on human coronavirus OC43, where Zanthoxylum bungeanum extract (Sichuan pepper) was found to inhibit CPE by impairing autolysosome accumulation and blocking virus-induced autophagic flux [16]. The authors demonstrated that the extract inhibited viral entry and caused an accumulation of LC3-II, a marker of autophagosomes, but also impaired the final degradation step (autolysosome formation), effectively trapping the virus in non-functional autophagic compartments. This suggests that OC43, and likely other betacoronaviruses, relies on intact autophagic flux for efficient replication, and its disruption is a potent anti-CPE strategy. This is a critical distinction from viruses that induce autophagy for replication versus those that require autophagic completion.
Cell-Cell Fusion as a Direct CPE Mechanism
While often a consequence of lytic infection, syncytia formation is itself a distinct and primary CPE mechanism for several virus families. It is driven by viral fusion proteins expressed on the surface of infected cells, which then interact with receptors on adjacent uninfected cells, leading to the merger of their plasma membranes. This process effectively bypasses the need for extracellular viral particle release for spread, enabling rapid viral dissemination and immune evasion.
The Paramyxoviridae and Herpesviridae families are masters of this strategy. As noted, NDV's F protein, in concert with the hemagglutinin-neuraminidase (HN) protein, drives syncytia formation in CEF and DF-1 cells [2]. The extensive multinucleated giant cells with large, fused nuclei are a direct consequence of this mechanism. Similarly, in Avian Influenza Virus (AIV), the CPE includes giant cell formation, though this is often accompanied by granularity and cell aggregation [2], reflecting the less pronounced fusion activity of the influenza hemagglutinin (HA) compared to paramyxovirus F proteins.
Among the herpesviruses, the Equine Herpesviruses (EHV), particularly EHV-1 and EHV-5, induce a characteristic CPE in RK-13 cells, visible 48-72 hours post-infection, which includes rounding and the formation of small foci of syncytia [5]. The fusion event is mediated by the viral glycoproteins gB, gH, and gL, and it is a key step in the virus's life cycle. For EHV-5, the CPE development was notably slower, often requiring up to 5 days and multiple passages to become evident [5]. This difference in CPE kinetics is a direct reflection of the molecular differences in replication kinetics and fusion protein activity between the alphaherpesvirus (EHV-1) and the gammaherpesviruses (EHV-2, EHV-5). The successful isolation of EHV-5, with its fastidious growth and slow CPE, is a notable achievement and underscores the importance of blind passaging for isolating slow-replicating cytopathic agents [5].
Replication Kinetics and the Temporal Profile of CPE
The rate at which CPE develops is a direct function of viral replication kinetics, specifically the speed at which the virus completes its replication cycle and the efficiency of its host-shutoff strategies. This is elegantly demonstrated by flaviviruses. A kinetic model based on real-time cell electric impedance (RTCA) technology compared West Nile Virus (WNV) and St. Louis Encephalitis Virus (SLEV) infection in Vero cells [8]. The time to 50% decrease in cell impedance (CIT50) was inversely proportional to the infectious dose and was significantly shorter for WNV than for SLEV. The analysis revealed that WNV's replication rate was approximately three times faster. This means that the accumulation of viral proteins and dsRNA, the subsequent activation of the cellular stress response (including PKR and RNase L pathways), and the induction of apoptosis occur more rapidly with WNV, leading to a faster and more aggressive CPE. This kinetic difference has practical implications for diagnostic workflows and for understanding the relative in vivo pathogenicity of these viruses [8, 22]. Similarly, clinical isolates of chikungunya virus (CHIKV) from India were categorized as "fast-growing" when CPE was observed within 24 hours post-infection in Vero cells [18]. These isolates were associated with a distinct acute cytokine profile (elevated IFN-alpha, IL-1RA, MCP-1), linking rapid in vitro CPE kinetics to a specific in vivo immunopathogenic signature [18].
The Host Factor: Cell-Type Specific Susceptibility and CPE Threshold
A critical determinant of CPE is not just the virus, but the host cell itself. The intrinsic susceptibility of a cell line dictates whether a productive, cytopathic infection will occur. This is vividly illustrated by Severe Fever with Thrombocytopenia Syndrome Virus (SFTSV) [6]. SFTSV infection induced clear CPE (rounding, detachment, plaque formation) in tiger (TLT), feline (CRFK, FCWF-4), and porcine (CPK) cell lines, while no CPE was observed in Vero E6, A549, BHK-21, or several deer and hedgehog-derived cell lines, despite detectable viral replication in some [6]. This demonstrates that the same viral genome can trigger a cytopathic cascade in one cellular background but not another. This is likely due to differences in the expression of viral receptors (e.g., DC-SIGN, NCAM for SFTSV), the integrity of the interferon signaling pathway (Vero cells are IFN-deficient, yet no CPE was seen, suggesting other blocks), and the activation status of downstream pro-apoptotic vs. pro-survival pathways. For example, the presence of a functional, virus-sensing RIG-I/MDA5 pathway in certain cells may trigger a rapid, protective IFN response that limits viral replication and CPE, whereas its absence or subversion in others allows unchecked replication and cytolysis [6].
Concluding Mechanistic Overview
The molecular pathogenesis of CPE is a multi-layered process where the virus's genetic blueprint (e.g., presence of a protease, a fusion protein, an interferon antagonist) interacts with the host cell's intrinsic machinery (apoptosis, autophagy, translation) to produce a visible phenotype. The examples detailed here--from the protease-driven host shutoff of PTV-5, the NS3-dependent apoptosis of cp BVDV, the fusion-mediated syncytia of NDV and EHV-1, the autophagic complex manipulations of PDCoV and OC43, to the kinetic and host-dictated differences seen in flaviviruses and SFTSV--underscore that CPE is not a single phenomenon but a diverse spectrum of cellular responses to a foreign invader. A deep, molecular-level understanding of these pathways is indispensable for the veterinary clinical pathologist, as it provides the framework for interpreting in vitro observations and connecting them to in vivo disease, ultimately guiding diagnostic strategies and the development of targeted antiviral interventions.
Classification and Morphological Characterization of CPE Types in Veterinary Viruses
The systematic classification of cytopathic effect (CPE) patterns constitutes a cornerstone of diagnostic veterinary virology, providing the first macroscopic evidence of viral replication in cell culture systems. As a clinical pathologist evaluating viral isolates, the ability to recognize, categorize, and interpret distinct CPE morphologies is essential for preliminary virus identification, guiding subsequent confirmatory testing, and understanding viral pathogenesis at the cellular level. This section provides an exhaustive analysis of the major CPE types observed in veterinary virology, correlating morphological patterns with specific viral families, underlying molecular mechanisms, and practical diagnostic implications.
The Diagnostic Significance of CPE Typing
Before the advent of molecular diagnostics, CPE observation was the primary method for detecting and characterizing viral agents. Even in the modern era of PCR and next-generation sequencing, CPE evaluation remains indispensable. The initial observation of a characteristic CPE in a susceptible cell line--whether it be rounding, syncytium formation, or inclusion body development--provides immediate, actionable information that can direct the diagnostic algorithm. For instance, the detection of syncytia in Vero cells inoculated with equine samples immediately raises suspicion for Equine Herpesvirus 1 or Equine Arteritis Virus, whereas the appearance of refractile, rounded cells in PK-15 cells inoculated with porcine samples suggests Porcine Teschovirus infection [1, 5]. This morphological triage is particularly valuable in resource-limited settings where molecular tools may be unavailable, and it remains a critical component of virus isolation protocols employed by reference laboratories worldwide, including those following WOAH (formerly OIE) guidelines.
Type I CPE: Cell Rounding and Detachment
The most frequently encountered CPE pattern in veterinary virology is cell rounding followed by progressive detachment from the monolayer. This morphological change reflects a profound disruption of the cellular cytoskeleton, often mediated by viral proteases that cleave intermediate filaments and actin microfilaments, or by the inhibition of host cell protein synthesis leading to cellular collapse.
Picornaviridae are archetypal inducers of this CPE type. Porcine Teschovirus, a member of the Picornaviridae family, induces characteristic rounding of PK-15 cells within 48-72 hours post-infection, with cells becoming highly refractile under phase-contrast microscopy before detaching in sheets [1]. The molecular basis involves the viral 2A protease-mediated cleavage of eukaryotic initiation factor 4G (eIF4G), shutting down cap-dependent host translation while allowing viral protein synthesis to proceed. Similarly, Foot-and-Mouth Disease Virus produces rapid and dramatic rounding of BHK-21 cells, with CPE evident as early as 24 hours post-inoculation, characterized by complete monolayer destruction within 48-72 hours [20]. The speed and completeness of this CPE correlate with the highly cytolytic nature of FMDV infection in epithelial cells.
Caliciviridae also produce rounding CPE, though often with distinct features. Feline Calicivirus induces rounding and detachment in feline cell lines such as CRFK, but the CPE is typically more focal and progresses more slowly than that of picornaviruses, with infected cells showing a characteristic "ballooning" degeneration before lysis [23]. This pattern reflects the virus's reliance on the host cell's translational machinery without the same degree of host shutoff seen in picornavirus infections.
Coronaviridae present a more nuanced rounding CPE. Porcine Epidemic Diarrhea Virus in Vero cells induces progressive rounding and syncytium formation, but the rounding is often preceded by cell-cell fusion, creating a mixed CPE pattern [14, 21]. The rounding component is driven by the disruption of the actin cytoskeleton mediated by the viral nonstructural proteins, particularly nsp1, which degrades host mRNA and induces cellular stress responses. Transmissible Gastroenteritis Virus produces a similar pattern in swine testicular cells, with rounding beginning at the periphery of the monolayer and progressing centripetally.
Type II CPE: Syncytium Formation (Cell Fusion)
Syncytium formation represents one of the most diagnostically useful CPE patterns, as it is relatively restricted to specific viral families and often correlates with the expression of viral fusion glycoproteins on the infected cell surface. The fusion of adjacent cell membranes creates multinucleated giant cells, which can be observed as large, irregularly shaped areas of the monolayer containing numerous nuclei arranged in a ring or cluster.
Paramyxoviridae are classic inducers of syncytial CPE. Newcastle Disease Virus produces prominent syncytia in chicken embryo fibroblast (CEF) and DF-1 cells, with fusion events beginning as early as 24-48 hours post-infection [3, 2]. The fusion process is mediated by the viral fusion (F) protein, which requires proteolytic cleavage by host cell proteases to become active. The extent of syncytium formation correlates directly with viral virulence; velogenic strains induce rapid, extensive fusion, while lentogenic strains produce minimal or no syncytia. Canine Distemper Virus similarly induces large, balloon-like syncytia in Vero cells expressing the SLAM receptor, with nuclei accumulating in the center of the fused mass [3].
Herpesviridae produce a distinctive syncytial CPE that is often slower to develop but highly characteristic. Equine Herpesvirus 1 in RK-13 cells induces focal areas of cell rounding that progress to small syncytia containing 3-10 nuclei, typically visible 48-72 hours post-inoculation [5]. The CPE is often described as "grape-like" clusters of rounded cells that eventually detach. Bovine Herpesvirus 1 produces similar syncytia in MDBK cells, though the fusion is often less extensive than that seen with paramyxoviruses. The molecular mechanism involves the viral glycoprotein gB, gD, and the gH/gL complex, which mediate membrane fusion during both entry and cell-to-cell spread.
Retroviridae can also induce syncytium formation, though this is typically a slower process. Feline Leukemia Virus and Bovine Leukemia Virus produce syncytia in permissive cell lines, but the CPE develops over 7-14 days and requires careful observation. The fusion is mediated by the viral envelope glycoprotein (Env) interacting with specific receptors on adjacent cells.
Type III CPE: Inclusion Body Formation
The formation of intracellular inclusion bodies--discrete, stainable structures within the nucleus or cytoplasm--represents a highly specific CPE type that can provide definitive diagnostic clues. These inclusions represent sites of viral replication, assembly, or the accumulation of viral proteins and nucleic acids.
Nuclear inclusions are characteristic of several DNA virus families. Canine Adenovirus (both types 1 and 2) produces large, basophilic intranuclear inclusions in canine kidney cells, which expand the nucleus and push chromatin to the periphery, creating a "Cowdry type A" inclusion body [2]. These inclusions are visible as early as 24-48 hours post-infection and are pathognomonic for adenovirus infection. Marek's Disease Virus, an alphaherpesvirus, produces intranuclear inclusions in chicken embryo fibroblast cells, though these are often smaller and more numerous than those seen with adenoviruses.
Cytoplasmic inclusions are prominent in several RNA virus families. Avian Influenza Virus produces eosinophilic cytoplasmic inclusions in DF-1 and CEF cells, which correspond to sites of viral ribonucleoprotein accumulation [2]. These inclusions are often perinuclear and may be accompanied by cell rounding and detachment. Rabies Lyssavirus produces the classic Negri bodies--eosinophilic, cytoplasmic inclusions in neurons and, in cell culture, in BHK-21 or Vero cells [12]. These inclusions are highly diagnostic but may require multiple blind passages before becoming apparent.
Poxviridae produce characteristic cytoplasmic inclusions known as B-type inclusions (Guarnieri bodies) or A-type inclusions depending on the virus. Fowl Pox Virus produces large, eosinophilic cytoplasmic inclusions in CEF cells, which are visible as early as 48 hours post-infection and are pathognomonic for avipoxvirus infection.
Cell-Type Dependency and CPE Variability
A critical concept in CPE classification is that the same virus can produce dramatically different CPE patterns depending on the cell line used. This phenomenon is well-illustrated by Severe Fever with Thrombocytopenia Syndrome Virus (SFTSV), which induces clear CPE in tiger-derived TLT cells and feline CRFK cells but fails to produce any visible CPE in Vero E6, A549, or BHK-21 cells, despite productive replication in some of these lines [6]. This cell-type dependency reflects differences in receptor expression, interferon signaling competence, and apoptotic pathway activation.
Similarly, Newcastle Disease Virus produces more rapid and extensive CPE in CEF cells compared to Vero cells, with syncytium formation occurring earlier and at higher titers in the avian cells [3, 2]. The Porcine Reproductive and Respiratory Syndrome Virus demonstrates a particularly striking cell-type dependency: it produces CPE in primary porcine alveolar macrophages (PAM) within 24-48 hours, characterized by rounding and detachment, but requires adaptation to produce CPE in MARC-145 cells, where the CPE is slower and less pronounced [19].
The Challenge of Differentiating CPE from Cytotoxicity
One of the most significant challenges in CPE-based diagnostics is distinguishing virus-induced CPE from chemical or physical cytotoxicity. This is particularly critical in disinfectant efficacy testing, where residual disinfectant can produce cellular changes that mimic viral CPE. A multi-step analytical framework has been developed to address this challenge, incorporating CPE observation, lactate dehydrogenase (LDH) release assays, MTT viability assays, and antigen-specific detection methods such as immunofluorescence and lateral flow immunoassays [10]. The key principle is that true viral CPE should correlate with the detection of viral antigens or nucleic acids, whereas cytotoxicity will be present in the absence of viral replication.
Quantification and Standardization of CPE Assessment
Modern approaches to CPE evaluation have moved beyond subjective observation to quantitative, automated methods. Real-time cell analysis (RTCA) systems based on cell-substrate electric impedance technology allow continuous monitoring of CPE development, generating kinetic parameters such as the CIT50 (time to 50% decrease in cell impedance) that correlate with viral infectious dose [8]. This technology has been applied to West Nile Virus and St. Louis Encephalitis Virus, demonstrating that CPE kinetics can differentiate between closely related flaviviruses based on their replication rates.
Automated imaging systems, such as the AID MultiSpot, enable whole-well scanning and quantification of CPE across multiple stages (1-4), providing objective, reproducible data that can be compared across experiments and laboratories [24]. These systems are particularly valuable for high-throughput screening of antiviral compounds, where CPE inhibition serves as the primary readout [13, 11, 9]. The resazurin-based CPE inhibition assay, which measures viable cell metabolism through the reduction of resazurin to fluorescent resorufin, has been validated for Feline Coronavirus and other veterinary viruses, providing a quantitative, objective endpoint that correlates well with viral replication [11].
Clinical Application and Diagnostic Performance of CPE in Viral Isolation and Identification
The observation of cytopathic effect (CPE) in cell culture remains a cornerstone of classical virology, serving as the primary phenotypic readout for virus isolation, identification, and characterization in diagnostic and research settings. Despite the increasing availability of molecular techniques, CPE-based virus isolation retains irreplaceable value for generating live virus stocks, conducting phenotypic antiviral susceptibility testing, and confirming the viability of viral pathogens in clinical specimens. As a veterinary clinical pathologist, I must emphasize that the diagnostic performance of CPE evaluation is fundamentally dependent upon the selection of appropriate cell lines, the recognition of virus-specific morphological patterns, and the rigorous differentiation of virus-induced cytopathology from cellular toxicity or senescence.
The Spectrum of CPE Morphologies Across Viral Families
The diagnostic utility of CPE observation lies in the fact that distinct virus families, and often individual virus species, produce characteristic cytopathic patterns that provide immediate preliminary identification clues. In the context of veterinary virology, the recognition of these patterns is essential for guiding confirmatory testing and interpreting isolation results.
Rounding and detachment represents the most commonly observed CPE pattern across numerous viral families. This morphology is characteristic of picornaviruses such as Porcine Teschovirus, where infection of PK-15 cells leads to progressive cellular rounding, shrinkage, and detachment from the monolayer, typically observable within 48-72 hours post-inoculation [1]. Similarly, Foot-and-Mouth Disease Virus induces rapid and pronounced rounding of BHK-21 cells, with CPE developing within 24-48 hours, enabling rapid presumptive diagnosis of this economically devastating pathogen [20]. The Bovine Viral Diarrhea Virus type 1a strain HN01 produces a classical cytopathic effect in MDBK cells characterized by cellular rounding and monolayer disruption, typically appearing after two passages and serving as a hallmark of the cytopathic biotype [4].
Syncytium formation is a hallmark of paramyxoviruses, herpesviruses, and certain retroviruses. Newcastle Disease Virus induces prominent cell fusion in both chicken embryo fibroblast (CEF) and DF-1 cells, resulting in the formation of multinucleated giant cells with intracytoplasmic inclusions, alongside cellular rounding and aggregation [2]. This syncytial pattern is also observed with Equine Herpesvirus 1 in RK-13 cells, where CPE typically appears 48-72 hours post-inoculation and progresses to involve the entire monolayer [5]. The diagnostic significance of syncytium formation cannot be overstated, as it immediately narrows the differential diagnosis and directs confirmatory testing toward specific viral families.
Focal degeneration and plaque formation is characteristic of many flaviviruses and orthomyxoviruses. West Nile Virus in Birds produces focal areas of cellular degeneration in Vero cells, progressing to complete monolayer destruction within 5-7 days [22]. The Avian Influenza Virus induces giant cell formation with intracytoplasmic inclusions and cellular aggregation, though the monolayer may remain partially intact, a pattern that distinguishes it from the more destructive CPE of paramyxoviruses [2]. The real-time monitoring of flavivirus-induced CPE using cell electric impedance technology has demonstrated that West Nile Virus exhibits more rapid cytolytic activity compared to St. Louis encephalitis virus, with mathematical modeling indicating approximately three-fold faster replication kinetics [8].
Cell Line Selection and Diagnostic Sensitivity
The diagnostic performance of CPE-based virus isolation is critically dependent upon the selection of permissive cell lines. Different viruses exhibit marked variations in their ability to replicate and produce CPE across different cell types, and the diagnostic laboratory must maintain a panel of cell lines optimized for the target pathogens.
Primary cells versus continuous cell lines represent a fundamental consideration. Primary CEF cells have demonstrated superior sensitivity for the isolation of Newcastle Disease Virus and Infectious Bursal Disease Virus, yielding higher infectious titers compared to the continuous DF-1 cell line [2]. However, for Avian Influenza Virus, DF-1 cells support higher viral replication, highlighting the virus-specific nature of cell line susceptibility [2]. The adaptation of viruses to continuous cell lines is a critical process for vaccine production, as demonstrated by the serial passage of Newcastle disease virus thermostable strain I-2 in CEF and Vero cells, where characteristic CPE appeared from passages 3 and 10, respectively, with progressive increases in infectious titer [3].
Species-specific cell lines are often essential for successful virus isolation. The isolation of Porcine Reproductive and Respiratory Syndrome Virus requires the use of porcine alveolar macrophages (PAM) or MARC-145 simian kidney cells, with CPE confirmation by immunofluorescence using specific monoclonal antibodies [19]. For aquatic viruses, the epithelioma papilloma cyprinid (EPC) cell line has proven invaluable for the isolation of Infectious Hematopoietic Necrosis Virus, where CPE inhibition assays have been used to evaluate antiviral compounds [25]. The Largemouth Bass Ranavirus produces characteristic CPE in EPC cells, with cellular rounding and detachment that can be quantitatively assessed using CCK-8 assays [26].
Novel cell lines for emerging pathogens continue to expand diagnostic capabilities. The development of tiger-derived TLT cells has provided a superior substrate for Severe Fever with Thrombocytopenia Syndrome Virus isolation, forming clear, easily countable plaques that facilitate accurate viral titration [6]. This contrasts with Vero E6 cells, which support high viral yields but fail to exhibit CPE, demonstrating that CPE induction is cell-type dependent and not solely a function of viral replication efficiency [6].
Differentiating CPE from Cytotoxicity: A Critical Diagnostic Challenge
One of the most significant challenges in CPE-based diagnostics is the accurate differentiation of virus-induced cytopathology from cellular toxicity caused by sample components, disinfectants, or other non-viral factors. This distinction is particularly critical in diagnostic settings where clinical specimens may contain cytotoxic substances.
Integrated analytical frameworks have been developed to address this challenge. In the evaluation of African Swine Fever Virus disinfectant efficacy, a multi-step approach combining CPE observation with lactate dehydrogenase (LDH) release assays, MTT viability assays, and antigen detection via lateral flow immunoassay and immunofluorescence has proven essential for distinguishing true viral CPE from disinfectant-induced cytotoxicity [10]. This framework revealed that virus-plus-disinfectant groups sometimes exhibited lower cytotoxicity than disinfectant-only groups, highlighting the complex interactions between viruses and chemical agents that can lead to misinterpretation of CPE results [10].
Quantitative approaches enhance the objectivity of CPE assessment. The use of resazurin-based CPE inhibition assays, which detect viable cells through their reduction of resazurin to fluorescent resorufin, provides a quantitative, objective readout that reduces observer bias [11]. This approach has been successfully applied to screen compounds against Feline Coronavirus and FIP, identifying chloroquine, mefloquine, and hexamethylene amiloride as inhibitors of virus-induced CPE at low micromolar concentrations [11]. Similarly, atomic force microscopy (AFM) has been employed to detect early virus-cell interactions by measuring changes in cell surface roughness, providing a novel method for determining neutralizing antibody titers without relying on subjective CPE observation [12].
Time-of-addition studies provide mechanistic insights that aid in CPE interpretation. The evaluation of antiviral compounds against Porcine Epidemic Diarrhea Virus using CPE reduction assays demonstrated that the timing of drug addition relative to infection critically influences the observed cytopathic effect, with early intervention producing the greatest protection [21]. This principle is equally applicable to diagnostic settings, where the timing of sample collection and processing can significantly impact CPE development and interpretation.
CPE in Neutralization Assays and Antiviral Screening
The application of CPE observation extends beyond virus isolation to include functional antibody detection and antiviral drug evaluation, where it serves as the primary readout for virus neutralization tests (VNT) and cytopathic effect inhibition assays.
Virus neutralization testing remains the gold standard for detecting functional neutralizing antibodies, despite the availability of ELISA-based methods. The Rift Valley Fever Virus VNT relies on the inhibition of CPE in Vero cells, with protection rates exceeding 97% at serum dilutions of 1:2-1:8 against challenge doses of 10^2-10^3 TCID50 [15]. The correlation between competitive ELISA and VNT is strong at high antibody concentrations but diverges at lower dilutions, emphasizing that CPE-based neutralization assays detect functional antibodies targeting viral glycoproteins, whereas ELISA detects binding antibodies against nucleocapsid proteins [15]. This functional distinction is critical for vaccine efficacy assessment and serological surveillance.
High-throughput screening for antiviral compounds has been revolutionized by CPE-based assays. The screening of an FDA-approved drug library against Canine Parvovirus using a CPE-based assay identified seven compounds with >50% CPE inhibition from 1,430 drugs tested, with nitazoxanide, closantel sodium, and closantel demonstrating dose-dependent inhibition confirmed by absolute PCR quantification and immunofluorescence [13]. The Feline Calicivirus CPE inhibition assay has been used to evaluate nineteen candidate compounds, identifying mefloquine as a potent inhibitor with additive effects when combined with recombinant feline interferon-omega [23].
Colorimetric CPE-based microneutralization assays provide objective, spectrophotometric readouts that enhance reproducibility and throughput. The evaluation of SARS-CoV-2 neutralizing antibodies using a colorimetric live virus micro-neutralization assay demonstrated that at least 50% of ELISA-positive samples are positive in neutralization, with the objective spectrophotometric method correlating well with subjective microscopic observation [9]. This approach has been adapted for veterinary applications, including the evaluation of Avian Influenza Virus neutralizing antibodies.
Quantitative CPE Assessment and Diagnostic Performance Metrics
The diagnostic performance of CPE-based methods is enhanced by quantitative approaches that provide objective, reproducible measurements of virus-induced cytopathology.
Real-time cell analysis (RTCA) using cell-substrate electric impedance technology enables continuous, label-free monitoring of CPE development. The application of RTCA to West Nile Virus and St. Louis encephalitis virus infection in Vero cells demonstrated that the time to 50% decrease in cell impedance (CIT50) is inversely proportional to virus infectious dose, providing a quantitative parameter for viral growth kinetics [8]. This technology also enables the quantification of neutralizing antibody activity, with the delay in CIT50 correlating well with plaque reduction neutralization test titers [8].
Automated CPE quantification using specialized readers reduces observer variability and enhances throughput. The AID MultiSpot system, combining an inverted microscope with a fluorescence EliSpot reader, enables CPE monitoring and quantification without cellular staining or medium removal, allowing continuous assessment of CPE progression [24]. The vSpot spectrum reader generates whole-well images from multiple plate formats, facilitating comparison between treatment groups and standardization of results across experiments [24].
TCID50 and plaque assay remain the standard methods for quantifying infectious virus, with CPE serving as the endpoint for both. The Infectious Salmon Anemia Virus and Viral Hemorrhagic Septicemia Virus are routinely titrated using CPE-based methods in appropriate fish cell lines. The Reed and Muench method for calculating TCID50 from CPE data remains widely used, with the accuracy of the calculation dependent upon the clarity and reproducibility of CPE endpoints [15].
Limitations and Interpretive Pitfalls
Despite its enduring value, CPE-based virus isolation and identification has significant limitations that must be recognized by the diagnostic pathologist.
Slow-growing or non-cytopathic viruses present particular challenges. Equine Herpesvirus 5 induces CPE only after 5 days of culture, often requiring multiple blind passages before cytopathic changes become apparent [5]. Similarly, Bovine Viral Diarrhea Virus non-cytopathic biotypes do not produce visible CPE, requiring detection by immuno
Correlation of CPE Patterns with Viral Pathogenicity and Virulence Determinants
The cytopathic effect (CPE) observed in cell culture is not merely a diagnostic epiphenomenon; it represents a direct, quantifiable in vitro correlate of the molecular interplay between viral virulence determinants and host cellular machinery. For the veterinary clinical pathologist, the pattern, kinetics, and severity of CPE provide critical insights into the pathogenic potential of an isolate, often predicting in vivo virulence with remarkable fidelity. This section delineates the mechanistic underpinnings of this correlation, drawing upon a diverse array of veterinary viral pathogens to illustrate how specific CPE signatures--ranging from syncytia formation to rapid lytic destruction--are inextricably linked to defined virulence factors.
Syncytia Formation as a Marker of Fusogenic Virulence
The induction of syncytia, or multinucleated giant cells, is a hallmark CPE associated with viruses that encode fusion glycoproteins capable of mediating cell-to-cell spread without release into the extracellular milieu. This pattern is particularly characteristic of paramyxoviruses and coronaviruses and is a direct reflection of the functional activity of their fusion machinery. For instance, Newcastle Disease Virus (NDV) is well-documented to cause "multicellular large nuclei, cell fusion and rounding of cells" in both chicken embryo fibroblast (CEF) and DF-1 cell lines [2]. The fusogenic capacity of NDV is directly attributable to the cleavage of the fusion (F) protein, a key virulence determinant. Strains with a multibasic cleavage site in the F protein, characteristic of velogenic (highly virulent) pathotypes, are activated by ubiquitous furin-like proteases, leading to extensive syncytia formation and rapid, disseminated CPE. In contrast, lentogenic (low virulence) strains possess a monobasic cleavage site requiring exogenous trypsin, resulting in restricted fusion and a less pronounced CPE. Thus, the extent of syncytia formation in vitro is a rapid, qualitative proxy for the molecular virulence of the NDV isolate.
Similarly, coronaviruses such as Porcine Epidemic Diarrhea Virus (PEDV) and Feline Coronavirus (FCoV) induce syncytia in susceptible cell lines, a process driven by the spike (S) protein. The S protein's S1/S2 cleavage site and the S2' fusion peptide are critical determinants of cell tropism and fusogenicity. In the context of FCoV, the emergence of a highly virulent biotype capable of causing feline infectious peritonitis (FIP) is associated with specific mutations in the S protein that enhance macrophage tropism and syncytia formation in vitro [11]. The CPE inhibition assay, using a resazurin-based readout, has been instrumental in screening for antiviral compounds against FCoV, where the protection from virus-induced CPE (i.e., inhibition of syncytia) directly correlates with reduced viral replication and, by extension, potential in vivo efficacy [11]. The correlation between fusogenic CPE and virulence is also evident in Porcine Deltacoronavirus (PDCoV), where infection of IPEC-J2 intestinal epithelial cells leads to characteristic CPE, and the virus's ability to cause systemic cytokine responses in gnotobiotic pigs is linked to its replicative fitness in enterocytes [7].
Lytic CPE and Host-Cell Shutoff: The Signature of Cytolytic Viruses
A rapid, lytic CPE characterized by cell rounding, detachment, and monolayer destruction is the classic signature of highly cytolytic viruses, particularly picornaviruses, flaviviruses, and certain asfarviruses. This pattern is a direct consequence of the virus's strategy to hijack the host cell's translational machinery, often through the cleavage of eukaryotic initiation factors (eIFs), leading to a "host-cell shutoff" that prioritizes viral protein synthesis at the expense of cellular integrity.
The isolation of a recombinant Porcine Teschovirus 5 (PTV-5) strain (SD2023) in PK-15 cells is a paradigmatic example. This strain induced "typical cytopathic effects" and was found to be a novel recombinant with a P1 gene derived from a PTV-5 minor parent and flanking regions from a PTV-4 major parent [1]. The CPE observed in vitro correlated with mild neurological symptoms and lymphocyte infiltration in the brain tissue of challenged piglets, confirming that the recombinant virus retained pathogenic potential [1]. The lytic CPE of PTV is a direct reflection of its picornaviral 2A and 3C proteases, which cleave eIF4G and other host factors, leading to the rapid demise of the infected cell. The speed and extent of this CPE can be used to gauge the relative virulence of different PTV genotypes, with more rapidly lytic strains often associated with more severe clinical outcomes.
Flaviviruses, such as West Nile Virus (WNV) and Chikungunya Virus (CHIKV), also exhibit a lytic CPE in Vero cells, but the kinetics can vary significantly between species and strains. Real-time cell analysis (RTCA) using cell electric impedance has provided a quantitative, high-resolution method to differentiate these kinetics. For instance, WNV-induced CPE in Vero cells was shown to be earlier and faster than that of St. Louis encephalitis virus (SLEV), with a mathematical model indicating that the replication rate of WNV was approximately three times faster [8]. This difference in CPE kinetics, quantified as the CIT50 (time to 50% decrease in cell impedance), directly correlated with the viral cytolytic activity. The RTCA system could quantify the level of cell protection by neutralizing antibodies, with the delay in CIT50 being well-correlated with antibody titer [8]. This demonstrates that the lytic CPE is not only a marker of virulence but also a functional readout for protective immunity. Similarly, clinical isolates of CHIKV from India were classified as "fast-growing" if they induced CPE within 24 hours post-infection in Vero cells, and these isolates were associated with elevated serum levels of specific pro-inflammatory cytokines (IFN-alpha, IL-1RA, IL-17F, MCP-1) in the acute phase of infection [18]. This underscores that the in vitro replication speed, as measured by CPE onset, is a surrogate marker for in vivo pathogenic potential and host immune activation.
The lytic CPE is also the cornerstone of virus titration and neutralization assays for highly pathogenic viruses like African Swine Fever Virus (ASFV). In disinfectant efficacy testing, a multi-step analytical framework was developed to differentiate ASFV-induced CPE from disinfectant-induced cytotoxicity, using LDH and MTT assays alongside CPE observation [10]. The ability of a disinfectant to inhibit CPE is the primary endpoint for determining virucidal efficacy, and the complexity of virus-disinfectant interactions (e.g., increased p30 signals after disinfection) highlights the need for careful interpretation of CPE data [10]. This framework is critical for ensuring that the observed CPE is truly virus-specific and not an artifact of chemical toxicity, a fundamental principle in any CPE-based assay.
Apoptosis and Non-Lytic CPE: The Subtle Pathogenesis of Persistent Viruses
Not all CPE is lytic. Many viruses, particularly parvoviruses and some retroviruses, induce apoptosis as a primary mechanism of cell death, leading to a more subtle, progressive CPE characterized by cell shrinkage, membrane blebbing, and nuclear condensation. This pattern is often associated with viruses that cause chronic or persistent infections.
Canine Parvovirus (CPV) is a classic example. In F81 cells, CPV infection leads to a characteristic CPE that is the basis for high-throughput screening of antiviral compounds [13]. The CPE is a direct consequence of the non-structural protein NS1, which induces cell cycle arrest and apoptosis. The study by Zhou et al. (2019) used a CPE-based high-throughput screening assay to identify FDA-approved drugs with anti-CPV activity, demonstrating that the inhibition of CPE (i.e., protection from apoptosis) correlated with a reduction in viral DNA replication and protein expression [13]. The antiapoptotic effects of one candidate drug, Nitazoxanide, were observed within 12 hours of treatment, regardless of CPV infection, indicating that the drug's mechanism of action involves modulating the host cell's apoptotic pathway [13]. This illustrates that the CPE pattern (apoptotic vs. lytic) can guide the selection of antiviral strategies, with antiapoptotic agents being potentially more effective against viruses that rely on this mechanism for cell death and viral release.
Similarly, Infectious Hematopoietic Necrosis Virus (IHNV), a rhabdovirus of salmonids, induces apoptosis in EPC cells. The imidazole arctigenin derivative EOA was shown to "significantly inhibit apoptosis induced by IHNV in EPC cells," which correlated with a reduction in CPE, viral titer, and ultimately, a 32.0% reduction in mortality in rainbow trout [25]. The study demonstrated that EOA's antiviral effect was most pronounced in the early stage of infection (1-4 days), and it repressed IHNV glycoprotein gene expression in target tissues (kidney and spleen) [25]. This direct link between the inhibition of apoptotic CPE in vitro and reduced mortality in vivo provides compelling evidence that the CPE pattern is a valid surrogate endpoint for evaluating the efficacy of antiviral therapeutics.
Non-Cytopathic Infections and the Concept of Viral Stealth
The absence of a visible CPE is, in itself, a critical piece of diagnostic and pathogenic information. Several important veterinary viruses, particularly pestiviruses and some gammaherpesviruses, can establish persistent infections without causing overt cell damage in vitro. This "stealth" phenotype is a key virulence determinant, as it allows the virus to evade host immune detection and establish lifelong infections.
Bovine Viral Diarrhea Virus (BVDV) exists as two biotypes: cytopathic (cp) and non-cytopathic (ncp). The cp biotype induces a characteristic CPE in MDBK cells, while the ncp biotype does not. The emergence of cp BVDV from a ncp progenitor through RNA recombination (e.g., insertion of host-derived sequences or duplication of viral sequences) is the molecular event that triggers the development of mucosal disease in persistently infected cattle. The study by Wang et al. (2014) isolated a BVDV 1a strain (HN01) that caused CPE in MDBK cells after two passages, and this cp strain was shown to be moderately pathogenic in calves, causing fever, leukopenia, and clinical signs [4]. The ability to induce CPE in vitro is therefore a direct marker of the cp biotype and its associated virulence in the context of a persistent ncp infection.
Similarly, Equine Herpesvirus 5 (EHV-5), a gammaherpesvirus, is known to "slowly induce a cytopathic effect in cell culture," and its isolation is not always successful [5]. In the study by Radalj et al. (2018), EHV-5 was isolated from only 6.1% of organ samples, and CPE was observed only on day 5 of the third passage [5]. This slow, subtle CPE is characteristic of a virus that establishes latency and causes chronic, progressive disease, such as equine multinodular pulmonary fibrosis (EMPF). The difficulty in isolating EHV-5 and the delayed CPE underscore the need for molecular detection methods (e.g., multiplex nested PCR) for accurate diagnosis [5]. The CPE pattern--or lack thereof--thus provides a critical clue to the virus's life cycle and pathogenic strategy.
Cell-Type Specificity of CPE and Host Range Determinants
The susceptibility of a given cell line to a virus and the resulting CPE pattern are not universal; they are governed by the presence of specific host cell receptors and intracellular factors. This cell-type specificity is a direct reflection of the virus's host range and tissue tropism in vivo.
The study of Severe Fever with Thrombocytopenia Syndrome Virus (SFTSV) in a panel of mammalian cell lines provides a striking example. SFTSV induced CPE in tiger-derived TLT cells, cat-derived CRFK and FCWF-4 cells, and porcine CPK cells, but not in Vero E6 (monkey), A549 (human), BHK-21 (hamster), deer-derived DFKT/DFLT, or hedgehog-derived HHoVT cells [6]. This differential CPE induction correlated with viral yield, with TLT, CRFK, and FCWF-4 cells producing high titers of infectious virus [6]. The authors noted that TLT cells formed "clear plaques that were easy to count," making them a superior cell line for plaque assays [6]. This study demonstrates that the ability of a virus to induce CPE is cell-type dependent and is a direct function of the compatibility between viral attachment proteins and host cell receptors. The choice of cell line for virus isolation and CPE observation must therefore be guided by the known host range of the virus, and the absence of CPE in a particular cell line does not preclude the virus's presence or pathogenicity in its natural host.
In the poultry industry, the adaptation of vaccine strains to different cell lines is a practical application of this principle. The NDV I-2 vaccine strain was adapted to both CEF and Vero cells, but the CPE was noted "beginning from passages 3 and 10" in CEF and Vero cells, respectively, indicating a differential adaptation kinetics [3]. The CEF-adapted virus induced higher antibody titers and provided 100% protection against challenge, while the Vero cell-adapted virus provided 95% protection [3]. This highlights that the CPE pattern during adaptation can be used to monitor the virus's fitness in a given cell substrate, which is critical for optimizing vaccine production. Similarly, Avian Influenza Virus (AIV) showed higher titers in DF-1 cells compared to CEF cells, while [Infectious Bursal Disease Virus](/knowledge/viruses/avian-viruses/in
Advanced Imaging and Quantitative Analysis Techniques for CPE Assessment
The traditional assessment of cytopathic effect (CPE) by subjective microscopic observation, while foundational to veterinary virology, is inherently limited by inter-operator variability, semi-quantitative ordinal scales, and an inability to capture the temporal dynamics of virus-induced cytopathology. The contemporary veterinary clinical pathologist now commands an arsenal of advanced imaging and quantitative analysis platforms that transform CPE evaluation from a descriptive art into a precise, data-rich science. These technologies enable high-throughput screening, kinetic profiling of viral replication, and the discrimination of true viral CPE from confounding phenomena such as chemical cytotoxicity--a distinction of paramount importance in antiviral drug discovery and disinfectant efficacy testing.
Real-Time Cell Electric Impedance Technology
Among the most transformative advances is the application of real-time cell analysis (RTCA) systems based on cell-substrate electric impedance. This technology, as elegantly demonstrated in flavivirus research, continuously monitors the electrical impedance across microelectrodes at the bottom of tissue culture wells, which is directly proportional to the number, adhesion, and morphology of adherent cells. When a virus such as West Nile Virus in Birds or St. Louis encephalitis virus infects a monolayer, the progressive CPE--cell rounding, detachment, and death--produces a quantifiable decrease in impedance over time [8]. The key kinetic parameter derived from this system is the CIT50, or the time required for a 50% reduction in cell impedance. A landmark study demonstrated that this parameter is inversely proportional to the input virus infectious dose, establishing RTCA as a tool for both viral titration and the quantitative assessment of neutralizing antibody-mediated protection [8]. Mathematical modeling of the impedance-based CPE kinetic curves revealed that the replication rate of West Nile Virus in Birds is approximately three times faster than that of St. Louis encephalitis virus, a finding that aligns with the known cytolytic activity of these flaviviruses [8]. The system proved exquisitely sensitive for quantifying the level of cell protection conferred by specific neutralizing antibodies; the delay in CIT50 correlated robustly with neutralizing antibody titers measured by conventional plaque reduction neutralization tests (PRNT) [8]. This approach eliminates the need for endpoint staining and provides continuous, label-free monitoring, making it ideal for kinetic studies of viruses that produce rapid and complete CPE, such as Avian Influenza Virus and Newcastle Disease Virus.
Automated Whole-Well Imaging and High-Content Analysis
The subjectivity inherent in microscopic CPE scoring has been addressed through the development of automated imaging platforms that provide whole-well quantification without the need for cellular staining or removal of culture medium. Instruments such as the AID MultiSpot and vSpot spectrum use inverted microscopy combined with EliSpot reader technology to scan entire wells at 4x to 20x magnification, generating comprehensive datasets that capture the spatiotemporal progression of CPE [24]. These systems allow researchers to correlate CPE stages (typically scored 1-4) with precise quantitative metrics, such as the percentage of confluent monolayer area exhibiting morphological changes, the number of syncytia per field, or the extent of cell detachment [24]. The ability to generate whole-well overviews ensures that rare but critical events--such as the formation of minute plaques by slowly replicating viruses--are not missed. This technology has proven particularly valuable in the assessment of African Swine Fever Virus disinfectant studies, where the ability to objectively differentiate disinfectant-induced cytotoxicity from virus-induced CPE is critical. In one study, Vero cell-adapted ASFV was evaluated using a multi-step analytical framework that integrated automated CPE imaging with lactate dehydrogenase (LDH) release assays, MTT metabolic activity assays, and antigen detection via lateral flow immunoassay and immunofluorescence [10]. The automated imaging component allowed precise quantification of CPE extent, revealing that disinfectant dilutions previously considered effective based on manual observation could, in fact, produce ambiguous results due to overlapping cytotoxic and cytopathic morphologies [10]. This underscores the necessity of objective, imaging-based CPE quantification in any regulatory or preclinical evaluation protocol.
Fluorescence-Based Reporter Assays and High-Throughput Screening
The field of antiviral drug discovery has been revolutionized by the marriage of CPE-based screening with fluorescent readouts, enabling miniaturization and automation. A classic example is the resazurin-based cytopathic effect inhibition assay, in which viable cells metabolically reduce the non-fluorescent substrate resazurin to the highly fluorescent resorufin. This assay was instrumental in identifying small molecule inhibitors of Feline Coronavirus and FIP, screening a panel of nineteen candidate compounds against feline coronavirus (FCoV) in Crandell-Rees feline kidney (CRFK) cells [11]. The assay detected CPE inhibition as a preservation of fluorescent signal, allowing calculation of half-maximal effective concentrations (EC50) with high throughput. The same approach was subsequently applied to identify inhibitors of Canine Parvovirus, where a CPE-based high-throughput screening assay was used to screen a library of 1,430 FDA-approved drugs, identifying seven compounds with greater than 50% CPE inhibition [13]. The three most potent candidates--Nitazoxanide, Closantel Sodium, and Closantel--were confirmed by absolute PCR quantification and indirect immunofluorescence assay (IFA), demonstrating that the primary CPE screen could reliably predict downstream antiviral activity [13]. Such fluorescence-based CPE inhibition assays have also been applied to evaluate the anti-coronavirus activity of natural products, including the oxazole-4-carboxamide derivative KB-2777, where CPE protection was corroborated by immunofluorescence detection of viral spike protein and TCID50 reduction [14].
Atomic Force Microscopy for Ultrastructural CPE Assessment
At the frontier of CPE imaging is atomic force microscopy (AFM), a technique that provides nanoscale resolution of cell surface topography without the need for fixation, labeling, or conductive coatings. In a groundbreaking application, AFM was used to quantify the neutralization activity of anti-rabies sera and immunoglobulin by measuring virus-induced changes in cell surface roughness [12]. The principle is elegant: rabies virus infection causes measurable alterations in the nanoscale topography of Vero and BHK-21 cell membranes--changes that are attenuated when the virus is pre-incubated with neutralizing antibodies. The assay titer is defined as the serum dilution that produces a 50% suppression of the increase in cell surface roughness [12]. This method reduces the time to obtain results from 21 days (as required by the mouse neutralization test) to just 2 days, while also adhering to the 3R principle (Replacement, Reduction, Refinement) by minimizing laboratory animal use [12]. The study demonstrated that AFM-based CPE assessment could reliably quantify neutralizing antibody titers within a range of 1:100 to 1:3000, with results consistent with those obtained by the mouse neutralization test when evaluated by Bland-Altman analysis [12]. This approach holds particular promise for viruses that produce subtle or non-lytic CPE, such as Rabies Lyssavirus, where conventional CPE scoring is often complicated by the slow and incomplete cytopathology characteristic of rhabdovirus replication.
Quantitative Multiparametric Frameworks for Differentiating CPE from Cytotoxicity
Perhaps the most clinically impactful application of advanced CPE quantification is in the differentiation of virus-induced CPE from confounding factors such as disinfectant cytotoxicity, compound toxicity, or culture condition artefacts. The work of Song and colleagues on African Swine Fever Virus disinfectant evaluation established a standardized framework integrating four complementary readouts: (1) subjective CPE observation, (2) LDH release (a marker of membrane integrity), (3) MTT reduction (a marker of mitochondrial metabolic activity), and (4) antigen-specific detection via lateral flow immunoassay (p30) and immunofluorescence (p54) [10]. This framework revealed a critical finding: that increased p30 antigen signals after disinfection and lower cytotoxicity in virus-plus-disinfectant groups compared to disinfectant-only groups indicate complex virus-disinfectant interactions that can be easily misinterpreted by CPE observation alone [10]. Such multiparametric approaches are now becoming the gold standard for evaluating disinfectants against enveloped viruses, and the veterinary clinical pathologist must be prepared to recommend them in diagnostic and regulatory contexts.
Similarly, in Porcine Teschovirus research, the induction of CPE in PK-15 cells was confirmed not only by light microscopy but also by integration with full-length genomic sequencing, recombination analysis, and quantitative assessment of viral RNA copy numbers by RT-qPCR [1]. This combination of imaging and molecular quantification provides a rigorous foundation for characterizing novel viral isolates. In Feline Calicivirus studies, the CPE inhibition assay was combined with virus yield reduction and plaque reduction assays to ensure that the observed cytoprotective effect of mefloquine was genuinely antiviral and not artefactual [23].
Digital Image Analysis for Plaque Assay Quantification
The classic plaque assay, while a cornerstone of virology, has also benefited from advanced imaging. The vSpot spectrum reader is capable of generating whole-well pictures from any plate format (6-384 well plates), allowing for automated plaque counting from crystal violet-stained or fluorescently labeled monolayers [24]. This eliminates the inter-operator variability inherent in manual plaque counting and enables the detection of micro-plaques that are invisible to the naked eye. In the context of Infectious Salmon Anemia Virus and other aquatic viruses, automated plaque quantification has become essential for standardizing viral titration across laboratories and for evaluating vaccine potency.
Integrating CPE Kinetics with Molecular and Serological Data
The true power of advanced CPE assessment lies in its integration with molecular and serological data streams. For example, in the evaluation of Rift Valley Fever Virus antibodies, the virus neutralization test (VNT) relies on the inhibition of CPE as its readout, but the interpretation of this endpoint is enhanced when CPE kinetic data are correlated with competitive ELISA results and TCID50 calculations [15]. A systematic study demonstrated that CPE protection in VNT was time- and dose-dependent, with high protection rates (>97%) at low serum dilutions against standard challenge doses, but that neutralizing efficacy decreased significantly at higher viral loads [15]. The divergence between binding antibody detection (cELISA) and functional neutralizing activity (VNT) at low antibody concentrations underscores the importance of CPE-based functional assays in confirming protective immunity [15].
Ultrastructural CPE Characterization by Electron Microscopy
Finally, for certain veterinary pathogens, the characterization of CPE at the ultrastructural level remains indispensable. In the study of Dengue virus (though not a veterinary pathogen per se, the methodological approach is transferable), transmission electron microscopy of Aedes albopictus C6/36 cell cultures allowed visualization of virus-induced morphological changes at the subcellular level, revealing the presence of virus particles associated with membrane structures and the nature of cellular degeneration [27]. For Avian Influenza Virus, giant cell formation with inclusions and aggregation of cells can be observed in both chicken embryo fibroblast (CEF) and DF-1 cells, and the quantification of syncytium size and number by image analysis provides a quantitative correlate of viral fusogenic activity [2].
Practical Considerations for the Veterinary Clinical Pathologist
The selection of an advanced CPE assessment platform must be guided by the specific virus, cell line, and experimental question. For high-throughput antiviral screening, fluorescence-based metabolic or live/dead assays in 96- or 384-well formats are preferred. For kinetic studies of virus replication and neutralization, RTCA systems provide unparalleled temporal resolution. For viruses that produce subtle or slow CPE, such as Equine Herpesvirus 5 or Aleutian Disease Virus, AFM or high-content imaging with machine learning-based morphological classification may be necessary. Regardless of the platform, the integration of CPE imaging with orthogonal assays--including viral RNA quantification by RT-qPCR, viral antigen detection by IFA, and assessment of cellular viability markers--remains the gold standard for rigorous CPE assessment. The veterinary clinical pathologist must also be cognizant of the limitations: the sensitivity of these techniques is influenced by cell density, passage number, and the inherent variability of primary cell cultures (such as porcine alveolar macrophages for Porcine Reproductive and Respiratory Syndrome Virus isolation) compared to continuous cell lines [19]. Standardization of protocols, including the use of positive and negative controls, is essential for reproducibility across laboratories and species.
The Road Ahead: Artificial Intelligence and Machine Learning
The next frontier in CPE assessment is the application of artificial intelligence (AI) and deep learning to automated image analysis. Convolutional neural networks trained on large datasets of CPE images could potentially classify CPE severity, identify virus-specific cytopathic signatures, and predict viral titer with greater accuracy than human observers. Preliminary studies have demonstrated that machine learning algorithms can differentiate between CPE caused by different virus families based on morphological features alone. As these technologies mature, the veterinary clinical pathologist will gain access to increasingly powerful tools for CPE quantification, enabling more precise diagnosis of viral infections, more efficient antiviral drug screening, and a deeper understanding of virus-host interactions at the cellular level.
Quality Control, Limitations, and Troubleshooting in CPE-Based Diagnostic Workflows
The cytopathic effect (CPE) remains a cornerstone of traditional virological diagnosis, yet its subjective nature and dependence on numerous biological and technical variables render it a potential source of diagnostic error. In the veterinary diagnostic laboratory, where pathogen identification carries immediate consequences for treatment, biosecurity, and trade, rigorous quality control (QC) measures are not merely advisable; they are essential. A sophisticated understanding of the inherent limitations of CPE-based workflows and a systematic approach to troubleshooting are required to maintain diagnostic accuracy and reproducibility. This section provides a comprehensive, pathologist-level analysis of the QC frameworks, the critical limitations that can compromise CPE interpretation, and the troubleshooting protocols necessary for a robust diagnostic workflow.
The Imperative for Standardized Quality Control
The foundational principle of QC in CPE-based diagnostics is the distinction between a true virus-induced effect and cellular damage from other causes. The most pervasive confounder in a diagnostic setting is cytotoxicity introduced by the sample itself. Clinical specimens--especially fecal homogenates, tissue macerates, and environmental swabs--often contain proteolytic enzymes, bacterial toxins, or residual disinfectants that can rapidly destroy a monolayer, mimicking viral CPE. This challenge is particularly acute in the evaluation of disinfectant efficacy, as demonstrated by Song et al. [10], who developed a multi-step analytical framework specifically to differentiate CPE from cytotoxicity when testing African Swine Fever Virus disinfectants. Their work showed that a 375x dilution of Virkon S achieved effective viral inactivation with significantly lower cytotoxicity, while the manufacturer-recommended 275x dilution caused considerable cell damage that could be misinterpreted as residual viral activity. The lesson is clear: any diagnostic laboratory performing virus isolation from field samples must include parallel toxicity controls--inoculating cell monolayers with the sample diluent alone or with heat-inactivated (56 degrees C for 30 min) sample aliquots--to establish a baseline for non-viral cellular damage.
A second pillar of QC is the cell line itself. Not all cell lines are equally susceptible to a given virus, and even within a single line, passage number, confluency, and metabolic state profoundly affect CPE kinetics. For Porcine Reproductive and Respiratory Syndrome Virus isolation, Ramos et al. [19] utilized both primary porcine alveolar macrophages (PAM) and the MARC-145 cell line, noting that while both supported viral replication, the CPE in PAM cells was more characteristic and appeared earlier, emphasizing the need for cell-line-specific QC acceptance criteria. Standard operating procedures (SOPs) must define acceptable passage ranges (e.g., passages 5-20 for Vero cells) and include routine mycoplasma testing, as contamination can suppress CPE development or induce cytopathic changes of its own [6]. The source of cells matters: primary cells like chicken embryo fibroblasts (CEF) may exhibit batch-to-batch variation, while continuous lines like DF-1 or Vero offer greater genetic uniformity but may have altered virus receptor expression over time [3, 2].
Limitations Inherent to Visual CPE Assessment
The most significant limitation of traditional CPE-based detection is its subjectivity and lack of quantitation. The standard practice of grading CPE on a 1+ to 4+ scale by microscopic observation is inherently operator-dependent. Two technicians viewing the same well may assign different scores, leading to inter-observer variability that is unacceptable in a regulatory or outbreak investigation context. The challenge of early detection is particularly acute; as noted by Preyer et al. [24], "in the beginning, only few cells show a CPE, which makes it difficult to estimate the success." This is where automated imaging systems, such as the AID MultiSpot or the xCELLigence real-time cell analysis (RTCA) platform, offer a transformative advantage. Fang et al. [8] demonstrated that RTCA-based monitoring of West Nile Virus in Birds infection in Vero cells generated a kinetic parameter, the CIT50 (time to 50% decrease in cell impedance), that was inversely proportional to infectious dose and provided an objective, quantitative readout far superior to subjective scoring. This technology eliminates the need for multiple sampling points and provides a continuous, operator-independent record of cell health.
Another profound limitation is the failure to detect non-cytopathic virus strains. Many viruses of critical veterinary importance can establish a persistent or latent infection without producing visible CPE. The definitive example is Bovine Viral Diarrhea Virus (BVDV): cytopathic (cp) biotypes cause classic CPE in MDBK cells, enabling isolation, while non-cytopathic (ncp) biotypes replicate silently, producing no visible change. Wang et al. [4] isolated a BVDV 1a strain (HN01) that only induced CPE after two blind passages in MDBK cells, underscoring that a single negative CPE reading cannot rule out infection. For ncp BVDV and other non-cytopathic viruses like Classical Swine Fever Virus, the diagnostic workflow must incorporate confirmatory methods--immunofluorescence (IFA), immunoperoxidase staining, or RT-PCR--on all samples, regardless of CPE status. The same principle applies to Equine Herpesvirus 5; Radalj et al. [5] found that EHV-5 isolates required up to five days and three blind passages in RK-13 cells before CPE appeared, highlighting the risk of false negatives if cultures are discarded prematurely.
The kinetics of CPE development also present a significant diagnostic limitation. Different viruses infect and destroy monolayers at vastly different rates. Fast-replicating viruses like Newcastle Disease Virus or Avian Influenza Virus can produce widespread CPE within 24-48 hours [3, 2], while others, such as Koi Herpesvirus or Salmonid Alphavirus, may require 5-7 days or multiple blind passages. If the laboratory terminates observation at a fixed, early time point (e.g., 72 hours), slow-growing viruses will be missed. The SOP must define a minimum observation period based on the target pathogen's known replication cycle, and more importantly, must require a minimum of two blind passages (typically 7 days each) before a sample is declared negative. This is especially critical for aquatic viruses; many, including Infectious Salmon Anemia Virus and Nervous Necrosis Virus, are notoriously slowly cytopathic in cell culture and require prolonged incubation.
Troubleshooting Common Failures in CPE-Based Workflows
When a clinical sample fails to produce CPE despite strong clinical and epidemiological suspicion of a viral etiology, a systematic troubleshooting approach is required. The most common causes for false-negative results fall into three categories: sample integrity, cell line susceptibility, and virus viability.
Sample integrity issues are the most frequent culprit. Improper storage (prolonged room temperature, repeated freeze-thaw cycles) leads to viral inactivation. The laboratory must enforce strict cold-chain protocols: samples should be collected into viral transport medium (VTM) with antibiotics, kept at 4 degrees C for no more than 24 hours, and stored at -80 degrees C for longer periods. The presence of neutralizing antibodies in a convalescent sample can also block viral entry and CPE development; this is a well-recognized phenomenon in Foot-and-Mouth Disease Virus isolation, where acute-phase samples are preferred [20]. If a sample is suspected to contain antibodies, it may be necessary to dilute it (e.g., 1:10 to 1:100) before inoculation to reduce antibody concentration.
Cell line selection is a critical troubleshooting variable. Many viruses exhibit a narrow host cell tropism. Porcine Deltacoronavirus, for instance, is notoriously difficult to isolate; Jung et al. [7] succeeded only after using porcine intestinal epithelial (IPEC-J2) cells, and even then, CPE was subtle and required IFA confirmation. If an initial isolation attempt using a standard cell line (e.g., Vero or PK-15) fails, the laboratory should attempt isolation in a more physiologically relevant cell line or in primary cells. For Epizootic Hematopoietic Necrosis Virus and Infectious Hematopoietic Necrosis Virus, epithelioma papulosum cyprinid (EPC) cells are the standard; attempts to grow these rhabdoviruses in mammalian cell lines will uniformly fail [26, 25].
If a sample does produce CPE, troubleshooting must confirm the specificity of the effect. Non-specific CPE can be induced by mycoplasma contamination, residual trypsin in the inoculum, or pH fluctuations in the medium. The laboratory should always include an uninfected control (cells receiving only medium) and a mock-infected control (cells receiving sterile sample diluent). If the mock-infected control develops CPE, the cell line or medium is contaminated, and all concurrent sample results are invalid. Confirmation of CPE as virus-specific requires a secondary assay. The immunofluorescence assay (IFA) is the gold standard for rapid confirmation; for example, Anggita et al. [6] used IFA to confirm Severe Fever with Thrombocytopenia Syndrome Virus infection in tiger-derived TLT cells, correlating CPE with viral antigen detection. Alternatively, a virus neutralization test (VNT) can be performed by mixing the CPE-positive supernatant with a specific antiserum; if the CPE is blocked, the virus is identified. Manenti et al. [9] demonstrated this principle for [SARS-CoV-2], but the methodology is directly transferable to animal viruses.
The Pitfalls of CPE in Antibody Neutralization Assays
CPE is not only used for virus detection but is the endpoint for the gold-standard virus neutralization test (VNT). Here, QC and troubleshooting are even more demanding. The VNT requires precise titration of the challenge virus to ensure a consistent, reproducible CPE in the control wells. If the challenge virus titer is too low, the neutralization readout will be falsely elevated; if too high, the serum will appear to have no neutralizing activity. Dafalla et al. [15] demonstrated this for Rift Valley Fever Virus, showing that high challenge doses (10^5 TCID50) overwhelmed the neutralizing capacity of even strongly positive sera, while lower doses (10^2-10^3 TCID50) allowed clear differentiation. The laboratory must standardize the challenge dose to 100-200 TCID50 per well and verify this titer by back-titration on each test day.
Another QC challenge in the VNT is the serum toxicity. Serum from some species, particularly pigs and horses, can be inherently cytotoxic to certain cell lines. Pre-heating the serum (56 degrees C, 30 min) inactivates complement and reduces non-specific toxicity, but some samples may still require dilution. If a serum control well (cells + serum alone) shows CPE, the serum is toxic, and the test result is invalid. The laboratory must also control for non-specific virus inhibition. Some sera contain lysozyme, defensins, or other innate immune factors that block virus entry in a non-specific, non-antibody-mediated manner. A truly specific neutralization should be confirmed by the absence of CPE only when specific antibody is present, not by general toxicity [15].
Instrumentation and Automated QC
The transition from subjective microscopy to automated, quantitative CPE assessment is the single most impactful advance for QC in this field. The xCELLigence RTCA system, as employed by Fang et al. [8], provides a continuous, impedance-based measurement of cell health that is far more sensitive than visual inspection. The CIT50 parameter (time to 50% decrease in impedance) provides an objective, operator-independent endpoint. This eliminates the variability of manual scoring and allows for precise comparison of virus isolates and antibody titers. For high-throughput screening, automated EliSpot readers (e.g., AID vSpot) can scan entire wells at multiple magnifications, quantifying CPE without the need for cell staining, thus allowing kinetic monitoring without disrupting the culture [24].
For plaque assays, the choice of stain is a QC variable. Crystal violet is inexpensive but only stains fixed cells and does not discriminate live from dead. The use of neutral red or MTT allows for live-cell staining and can be read spectrophotometrically, providing a quantitative endpoint that correlates with viral titer. Song et al. [10] incorporated both LDH release and MTT reduction assays into their ASFV disinfectant framework, demonstrating that MTT-based viability assessment provided a more sensitive and objective measure of CPE than visual inspection alone.
Addressing the Specific Challenges of Aquatic Virology
CPE-based diagnostics in aquatic species present unique QC challenges. Many aquatic viruses, particularly those infecting crustaceans and mollusks, are difficult or impossible to grow in continuous cell lines. For White Spot Syndrome Virus and Yellow Head Virus, there is no established continuous cell line; diagnosis relies on PCR or histopathology, with CPE-based isolation playing no role. However, for finfish viruses, cell culture remains essential. The temperature of incubation is a critical variable. Infectious Pancreatic Necrosis Virus replicates optimally at 15-18 degrees C, while Channel Catfish Virus requires 25-30 degrees C. The laboratory must maintain dedicated incubators for different temperature regimens and rigorously monitor temperatures using continuous logging devices.
A final, subtle QC issue is the serum batch effect in cell culture medium. Fetal bovine serum (FBS) can contain bovine viral antibodies (e.g., anti-BVDV) that neutralize contaminating viruses or inhibit the growth of target viruses. For the isolation of Porcine Epidemic Diarrhea Virus, Sirichokchatchawan et al. [21] emphasized the use of FBS that had been heat-inactivated and certified free of anti-coronavirus antibodies. Standard practice in any virology laboratory should be to screen FBS batches for the presence of antibodies to common bovine viruses and to use gamma-irradiated or certified virus-free serum for all diagnostic work.
The pathologist overseeing a CPE-based diagnostic workflow must be vigilant, systematic, and aware of the myriad ways in which biological variability and technical artifacts can compromise results. Rigorous QC--including toxicity controls, cell-line characterization, and the incorporation of confirmatory molecular or immunological assays--is non-negotiable. The transition to automated, impedance-based or imaging-based CPE quantification offers a path to standardized, objective, and reproducible diagnostics that meet the highest standards of veterinary clinical pathology.
References
[1] Ye K, Sun H, Ma Z, Sun Y, Zhang L, Bai J, et al.. Molecular characterization and pathogenicity of a porcine Teschovirus 5 isolate in Shandong Province in China.. Veterinary Microbiology. 2026. DOI: https://doi.org/10.1016/j.vetmic.2026.110989
[2] Anam S, Rahman S, Ali S, Saeed M, Goyal S. Comparative growth kinetic study of Newcastle disease virus, infectious bursal disease virus and avian influenza virus in chicken embryo fibroblast and DF-1 cell lines.. Polish journal of veterinary sciences. 2021. DOI: https://doi.org/10.24425/pjvs.2021.137664
[3] Yami MM, Akalu M, Bayisa B, Tefera TA, Hurisa TT, Ayele G, et al.. Evaluation of vero and chicken embryo fibroblast cell-based newcastle disease vaccine efficacy in chickens. Poultry Science. 2025. DOI: https://doi.org/10.1016/j.psj.2025.105624
[4] Wang W, Shi X, Tong Q, Wu Y, Xia M, Ji Y, et al.. A bovine viral diarrhea virus type 1a strain in China: isolation, identification, and experimental infection in calves. Virology Journal. 2014. DOI: https://doi.org/10.1186/1743-422X-11-8
[5] Radalj A, Nišavić J, Krnjaić D, Valcic M, Jovanović T, Veljović L, et al.. Detection and molecular characterization of equine herpesviruses 1, 2, and 5 in horses in the Republic of Serbia. Acta Veterinaria Brno. 2018. DOI: https://doi.org/10.2754/AVB201887010027
[6] Anggita M, Nyampong S, Hu W, Shimoda H, Hayasaka D. Different Susceptibility of Mammalian Cell Lines to Severe Fever with Thrombocytopenia Syndrome Virus Infection. Viruses. 2025. DOI: https://doi.org/10.3390/v17101380
[7] Jung K, Miyazaki A, Hu H, Saif L. Susceptibility of porcine IPEC-J2 intestinal epithelial cells to infection with porcine deltacoronavirus (PDCoV) and serum cytokine responses of gnotobiotic pigs to acute infection with IPEC-J2 cell culture-passaged PDCoV. Veterinary Microbiology. 2018. DOI: https://doi.org/10.1016/j.vetmic.2018.05.019
[8] Fang Y, Ye P, Wang X, Xu X, Reisen W. Real-time monitoring of flavivirus induced cytopathogenesis using cell electric impedance technology. Journal of Virological Methods. 2011. DOI: https://doi.org/10.1016/j.jviromet.2011.02.013
[9] Manenti A, Maggetti M, Casa E, Martinuzzi D, Torelli A, Trombetta C, et al.. Evaluation of SARS-CoV-2 neutralizing antibodies using a CPE-based colorimetric live virus micro-neutralization assay in human serum samples. Journal of Medical Virology. 2020. DOI: https://doi.org/10.1002/jmv.25986
[10] Song S, Shin K, Kim S, Joo YY, Han B, Park S, et al.. A Practical Framework for ASFV Disinfectant Evaluation: Differentiating Cytopathic Effects from Cytotoxicity via Integrated Analytical Methods. Pathogens. 2025. DOI: https://doi.org/10.3390/pathogens14050451
[11] Identification and characterisation of small molecule inhibitors of feline coronavirus replication. Veterinary Microbiology. 2014. DOI: https://doi.org/10.1016/j.vetmic.2014.10.030
[12] Generalov S, Erokhin PS, Kuznetsov OS, Abramova EG, Zhulidov IM, Osina NA. Determining the Specific Activity of Anti-Rabies Sera and Immunoglobulin Using Atomic Force Microscopy of Cell Cultures. Avicenna journal of medical biotechnology. 2021. DOI: https://doi.org/10.18502/ajmb.v13i3.6362
[13] Zhou H, Su X, Lin L, Zhang J, Qi Q, Guo F, et al.. Inhibitory Effects of Antiviral Drug Candidates on Canine Parvovirus in F81 cells. Viruses. 2019. DOI: https://doi.org/10.3390/v11080742
[14] Jung B, Na W, Yeom M, Lim J, Do H, Jang G, et al.. Screening-Identified Oxazole-4-Carboxamide KB-2777 Exhibits In Vitro Anti-Coronavirus Activity. Pharmaceutics. 2025. DOI: https://doi.org/10.3390/pharmaceutics17111477
[15] Dafalla O, Alashor AA, Hussien M, Noureldin E, Ageep T, Najmi M, et al.. Integrated Laboratory Evaluation of Rift Valley Fever Virus Antibodies Using the Competitive ELISA and Virus Neutralization Test. Pathogens. 2026. DOI: https://doi.org/10.3390/pathogens15030264
[16] Park H, Ha IJ, Jin Y, Kwon S. Sichuan pepper, Zanthoxylum bungeanum Maxim., suppresses human coronavirus OC43 infection by inhibiting viral entry and impairing autolysosome accumulation. Chinese Medicine. 2026. DOI: https://doi.org/10.1186/s13020-026-01388-y
[17] Habib M, Jin Y, Kim Y, Min J, Ha I, Lee S, et al.. Anticoronavirus activity of Rhizome of Dryopteris crassirhizoma via multistage targeting of virus entry and viral proteases, Mpro and PLpro.. Journal of Ethnopharmacology. 2024. DOI: https://doi.org/10.1016/j.jep.2024.118490
[18] Babu N, Mahilkar S, Jayaram A, Ibemgbo S, Mathur G, Shetty U, et al.. Cytokine profile, neutralisation potential and viral replication dynamics in sera of chikungunya patients in India: a cross-sectional study. The Lancet Regional Health - Southeast Asia. 2023. DOI: https://doi.org/10.1016/j.lansea.2023.100269
[19] Ramos N, Mirazo S, Castro G, Cabrera K, Osorio F, Arbiza J. First-time detection of porcine reproductive and respiratory syndrome virus (PRRSV) infection in Uruguay.. Transboundary and Emerging Diseases. 2018. DOI: https://doi.org/10.1111/tbed.12813
[20] Muchie BT, Wubshet A, Haile A, Deyo HM, Tarasiuk K, Pejsak Z, et al.. Isolation and Molecular Identification of Foot and Mouth Disease Virus Circulating Around Central Ethiopia.. 2021. DOI: https://doi.org/10.21203/RS.3.RS-436692/V1
[21] Sirichokchatchawan W, Temeeyasen G, Nilubol D, Prapasarakul N. Protective Effects of Cell-Free Supernatant and Live Lactic Acid Bacteria Isolated from Thai Pigs Against a Pandemic Strain of Porcine Epidemic Diarrhea Virus. Probiotics and Antimicrobial Proteins. 2017. DOI: https://doi.org/10.1007/s12602-017-9281-y
[22] Hady HA, Yossef NM, El-Barrawy M, Hashish M, El-Ela NA. West Nile Virus Existence in Humans and Mosquitoes in Alexandria Governorate.. 2007. DOI: https://doi.org/10.21608/jhiph.2007.22574
[23] McDonagh P, Sheehy P, Fawcett A, Norris J. Antiviral effect of mefloquine on feline calicivirus in vitro. Veterinary Microbiology. 2015. DOI: https://doi.org/10.1016/j.vetmic.2015.02.007
[24] Preyer R, Kevin S, Gladyschuk V, Klimovich A, Ettischer-Schmid N. Solutions for quantifying cytopathic effects after viral infection in vitro. Journal of Immunology. 2024. DOI: https://doi.org/10.4049/jimmunol.212.supp.0856.6474
[25] Hu Y, Chen W, Shen Y, Zhu B, Wang G. Synthesis and antiviral activity of a new arctigenin derivative against IHNV in vitro and in vivo.. Fish and Shellfish Immunology. 2019. DOI: https://doi.org/10.1016/j.fsi.2019.07.006
[26] Chen M, Xue M, Zhou Y, Xu C, Jiang N, Fan Y, et al.. Antiviral effects of rhein on largemouth bass ranavirus (LMBRaV).. Fish and Shellfish Immunology. 2024. DOI: https://doi.org/10.1016/j.fsi.2024.110019
[27] Nogueira R, Schatzmayr H, Miagostovich M, Farias MDF, Filho JDCO. Virological study of a dengue type 1 epidemic at Rio de Janeiro.. Memórias do Instituto Oswaldo Cruz. 1988. DOI: https://doi.org/10.1590/S0074-02761988000200012 *** Disclaimer: This article is for educational and informational purposes only. It is not intended to substitute for professional veterinary advice, diagnosis, treatment, or regulatory guidance. Always consult a licensed veterinarian or qualified specialist regarding animal health, disease diagnosis, and therapeutic decisions.