
Trichomonosis in Wild Birds: Pathogenesis, Diagnostic Techniques, and Conservation Impact
Introduction
Trichomonosis is a parasitic disease of wild birds caused primarily by the flagellate protozoan Trichomonas gallinae. This infection, often termed "canker" in pigeons and doves and "frounce" in raptors, has emerged as a significant conservation threat for several avian species, particularly granivorous finches (Fringillidae) and columbids (Columbidae). The etiologic agent is a single-celled, anaerobic, flagellated protozoan belonging to the order Trichomonadida. Although several Trichomonas species have been described in birds, T. gallinae is the most commonly identified pathogen, responsible for epidemic outbreaks with high morbidity and mortality [1, 2]. In recent decades, trichomonosis has expanded its host range, causing dramatic population declines in European greenfinches (Chloris chloris) and chaffinches (Fringilla coelebs), and has been linked to the decline of the endangered pink pigeon (Nesoenas mayeri) on Mauritius [3, 4]. This review provides an exhaustive examination of the pathogenesis, diagnostic techniques, and conservation impact of trichomonosis in wild bird populations, emphasizing the biophysical and molecular mechanisms underlying host-parasite interactions that drive epidemic dynamics.
Pathogenesis
Parasite Biology and Transmission
Trichomonas gallinae is an extracellular, flagellated protozoan that replicates asexually by longitudinal binary fission. The trophozoite, the only life stage identified in avian hosts, is ovoid to pyriform, measuring 5-15 µm in length, and possesses three anterior flagella and a posterior flagellum that forms an undulating membrane [5]. The organism lacks mitochondria and relies on hydrogenosomes for anaerobic energy metabolism, making it highly dependent on carbohydrate-rich environments such as the upper digestive tract mucosa [6]. Transmission occurs primarily through direct oral contact, contaminated feed or water, and during courtship feeding (particularly in pigeons via crop milk) [7, 8]. Raptors acquire infection by consuming infected prey, linking predator-prey dynamics to disease spread [9].
Host-Pathogen Interactions
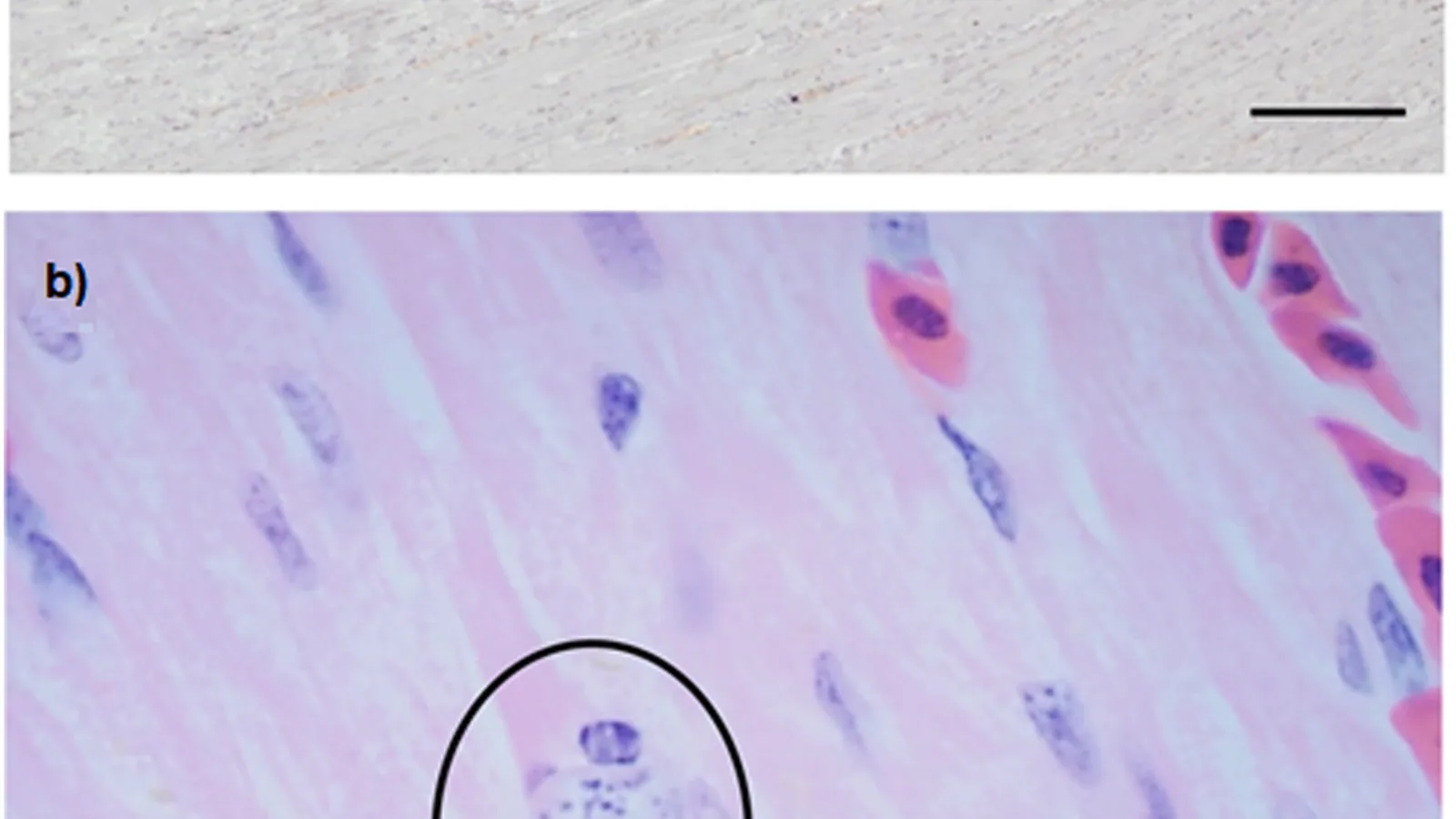
The pathogenesis of trichomonosis is driven by parasite adherence to host mucosal epithelium and subsequent secretion of proteolytic enzymes, including cysteine proteases, that degrade extracellular matrix components and facilitate tissue invasion [10, 11]. The trophozoites colonize the oropharynx, esophagus, and crop, causing caseous necrotic lesions that can obstruct the gastrointestinal tract. Histopathologically, lesions are characterized by intense lymphoplasmacytic and heterophilic inflammation with extensive fibrin deposition and necrosis [12]. The parasite's virulence is associated with strain-specific expression of surface adhesins and the ability to modulate host immune responses through immunoglobulin cleavage and apoptosis of macrophages [13, 14]. In finches, disease progression is remarkably rapid, with death often occurring within 4-7 days post-infection, whereas columbids may harbor subclinical infections for weeks [15].
Clinical Manifestations in Different Avian Orders
Clinical signs vary by species and age. In Columbiformes (pigeons and doves), caseous yellow plaques are commonly observed on the palate, esophagus, and crop. Infected birds show weight loss, regurgitation, excessive salivation, and dysphagia [16]. In Passeriformes (finches, sparrows), the disease manifests as a swelling of the neck region due to necrotic inflammation of the crop and esophagus, often visible externally as a "bulging neck" [17]. Raptors infected through prey ingestion may develop similar lesions but often present with emaciation, lethargy, and respiratory distress due to extension of lesions into the trachea [18]. Subclinical carriers are epidemiologically significant; they shed parasites intermittently in feces and saliva, maintaining transmission cycles even during non-epidemic periods [19]. Coinfections with other pathogens, such as those described in the Feline Upper Respiratory Tract Infection Complex: Multiplex PCR Panel Interpretation and Treatment Algorithms, can exacerbate clinical outcomes, though this has not been extensively studied in wild birds.
Molecular Mechanisms of Tissue Damage
The cysteine proteases of T. gallinae (e.g., TgCP-1) degrade host immunoglobulins, fibronectin, and collagen, promoting tissue necrosis and immune evasion [20]. Additionally, the parasite induces apoptosis in avian macrophages via caspase-dependent pathways, reducing the host's ability to contain infection [21]. Transcriptomic analyses of infected avian tissues reveal upregulation of genes encoding pro-inflammatory cytokines (IL-1β, IL-6) and downregulation of mucosal barrier proteins [22]. These molecular changes correlate with the severity of lesion formation and systemic metabolic disturbances, including hypoglycemia and dehydration [23].
Diagnostic Techniques
Accurate and timely diagnosis is critical for outbreak management and conservation interventions. Diagnostic approaches range from field-deployable microscopic examination to molecular and serological assays. The table below compares the principal methods used in wild bird trichomonosis diagnostics.
| Diagnostic Method | Target | Sensitivity | Specificity | Turnaround Time | Field Applicability |
|---|---|---|---|---|---|
| Wet mount microscopy | Trophozoites | Moderate (50-70%) | High (if experienced) | 10-15 minutes | Excellent |
| InPouch culture | Viable organisms | High (80-95%) | High | 2-5 days | Good (requires incubator) |
| Conventional PCR (18S rRNA, ITS1/2) | DNA | High (90-98%) | High | 4-6 hours | Limited (needs lab) |
| Quantitative real-time PCR (qPCR) | DNA | Very high (>98%) | Very high | 2-4 hours | Limited |
| Enzyme-linked immunosorbent assay (ELISA) | Serum antibodies | Moderate (60-80%) | Moderate | 3-4 hours | Moderate |
Microscopy and InPouch Culture
Direct microscopic examination of wet mounts from oral or crop swabs is the simplest diagnostic method. A saline-moistened swab is rolled onto a glass slide, covered with a coverslip, and examined at 200-400× magnification for flagellated trophozoites exhibiting jerky, rolling motility [24]. Sensitivity is limited in low-burden infections and when samples have been frozen or refrigerated. The InPouch culture system (a self-contained plastic bag with a liquid medium) has become the gold standard for field diagnosis in wild bird studies. A swab is inserted into the pouch, and after incubation at 37°C for 2-5 days, the presence of motile trichomonads is confirmed microscopically [25, 26]. This method allows semi-quantitative estimation of parasitic load and is particularly useful for detecting subclinical carriers. However, culture can be compromised by bacterial overgrowth and requires a portable incubator, which may be logistically challenging in remote field settings.
Molecular Diagnostics
Polymerase chain reaction (PCR) targeting the internal transcribed spacer (ITS) regions of ribosomal DNA (ITS1-5.8S-ITS2) provides species-level identification and can differentiate T. gallinae from closely related species such as T. stableri and Tetratrichomonas spp. [27, 28]. Conventional PCR followed by Sanger sequencing of the ITS region is widely used for phylogenetic analysis and outbreak tracking. Quantitative real-time PCR (qPCR) offers increased sensitivity and allows quantification of parasite DNA, which correlates with lesion severity [29]. Multiplex PCR panels, analogous to those used for respiratory pathogens in companion animals (see Feline Upper Respiratory Tract Infection Complex: Multiplex PCR Panel Interpretation and Treatment Algorithms), have been developed for simultaneous detection of Trichomonas spp. and other avian pathogens [30]. Molecular methods are essential for confirming infection in decomposed carcasses and for detecting mixed infections with multiple trichomonad species [31].
Serological Techniques
Enzyme-linked immunosorbent assays (ELISA) detecting anti-T. gallinae antibodies have been used in captive pigeon populations but show limited sensitivity in wild birds due to rapid disease progression before seroconversion [32]. Western blot analysis of serum proteins reveals specific bands corresponding to parasite surface antigens (e.g., 50 kDa adhesin), but this technique is not field-deployable and is primarily used for epidemiological surveys [33]. Point-of-care serological tests, similar to those described for Feline Leukemia Virus (FeLV) and Feline Immunodeficiency Virus (FIV): Point-of-Care Testing and Clinical Management, have not been developed for trichomonosis in birds, representing a gap in rapid diagnostic capability.
Diagnostic Workflow
The following Mermaid diagram outlines a recommended diagnostic algorithm for wild bird trichomonosis surveillance and outbreak investigation.
flowchart TD
A[Clinically suspect bird or carcass] --> B[Oropharyngeal/crop swab]
B --> C{Field conditions?}
C -->|Basic| D[Wet mount microscopy immediate]
C -->|Moderate| E[InPouch culture at ambient temperature]
C -->|Advanced| F[Swab stored in DNA/RNA Shield]
D --> G["Positive: trophozoites seen"]
D --> H["Negative: no motility"]
E --> I[Incubate 37°C x 72h]
I --> J[Microscopic examination]
J --> K["Positive: motile trichomonads"]
J --> L["Negative: no growth"]
F --> M[Laboratory PCR]
M --> N[ITS1-5.8S-ITS2 conventional PCR]
N --> O[Amplicon sequencing for species ID]
M --> P[Real-time qPCR for quantification]
L --> Q[Consider alternative diagnosis or PCR from culture]
H --> R[PCR directly from swab DNA]
R --> S["Result: confirmed positive/negative"]
K --> T[Epidemiological data collection]
O --> T
P --> T
S --> T
Conservation Impact
Epidemic Outbreaks in Finches and Pigeons
The most notable trichomonosis epidemics in wild birds have occurred in European passerines, particularly greenfinches and chaffinches. Starting in the mid-2000s, reports of mass mortality emerged from the United Kingdom, with annual peaks in late summer and autumn [34]. The disease spread rapidly across continental Europe, causing population declines of 30-50% in some regions [35]. Epidemic dynamics are driven by high bird densities at garden feeding stations, where contaminated food and perching surfaces facilitate horizontal transmission [36]. Behavioral factors such as flocking and communal feeding increase the basic reproduction number (R0) above 1, sustaining outbreaks across seasons.
In columbids, trichomonosis is enzootic but can produce epizootics when immune-naive populations are exposed to highly virulent strains. The endangered pink pigeon of Mauritius suffered severe declines in the 1990s due to trichomonosis, with mortality rates exceeding 50% in some years; conservation interventions including supplementation with oral antiprotozoal agents (ronidazole) and management of feeding stations helped stabilize the population [37, 38]. Similarly, band-tailed pigeons (Patagioenas fasciata) in North America have experienced periodic die-offs linked to T. gallinae infection, often exacerbated by concurrent infections with avian poxvirus or circovirus (see Pigeon Circovirus and Young Pigeon Disease) [39, 40].
Population-Level Consequences
Long-term monitoring data from the United Kingdom and the Netherlands indicate that trichomonosis has caused persistent reductions in greenfinch breeding populations, with recovery hindered by continued transmission at supplementary feeding sites [41]. Modeling studies suggest that seasonal outbreaks in finches are sustained by carrier individuals and that population recovery requires a reduction in pathogen prevalence below a threshold of 10-15% [42]. The disease also impacts raptor populations indirectly: when prey species decline, raptors may switch to alternative prey that are less competent hosts, but this behavioral shift can increase exposure to other pathogens [43]. In some ecosystems, trichomonosis acts as a density-dependent regulating factor, but when combined with habitat loss or climate stress, it can push small populations toward local extinction.
Interspecific Transmission and Host Range
T. gallinae exhibits a broad host range among bird orders. In addition to Columbiformes, Passeriformes, and Falconiformes, it has been documented in Galliformes, Psittaciformes, and Strigiformes [44, 45]. Strain typing using ITS sequencing has identified specific genotypes associated with finch outbreaks (e.g., A1 genotype) that are distinct from those circulating in pigeons, suggesting some degree of host adaptation [46]. However, transmission between columbids and passerines does occur, particularly when shared water sources are contaminated. The emergence of finch trichomonosis has been attributed to a host shift from pigeons, likely facilitated by the increasing overlap of these species at anthropogenic feeding sites [47]. This pattern echoes the cross-species transmission dynamics seen in other wildlife diseases, such as Avian Influenza A(H5N1) in Poultry and Wild Birds: Current Epidemiology, Molecular Diagnostics, and Biosecurity, where bridging hosts facilitate pathogen spillover between wild and domestic populations.
Mitigation Strategies and Conservation Recommendations
Management of trichomonosis in wild bird populations focuses on reducing transmission risk at artificial feeding stations. Key recommendations include regular cleaning of feeders with a 10% bleach solution, rotating feeder locations to prevent accumulation of contaminated feces, and temporarily removing feeders during outbreak peaks [48]. In captive breeding programs for endangered columbids, prophylactic treatment with metronidazole or ronidazole in feed has proven effective, though concerns about development of resistance and potential toxicity limit long-term use [49]. Surveillance programs combining molecular diagnostics with citizen science reporting (e.g., garden birdwatch schemes) have enabled rapid detection of outbreaks and targeted interventions [50]. Further research is needed to develop vaccines for wild birds, as current efforts are limited to experimental trials in pigeons.
Conclusion
Trichomonosis caused by Trichomonas gallinae represents a major parasitic threat to wild bird biodiversity, particularly for finches and pigeons. The pathogenesis involves rapid mucosal invasion, necrosis, and immune modulation by flagellate trophozoites. Diagnostic capabilities have advanced from simple wet mount microscopy to InPouch culture and highly sensitive molecular methods such as PCR and qPCR, enabling accurate species identification and quantification of parasite loads. Conservation impact is profound, with epidemic outbreaks driving population declines and threatening vulnerable island endemics. Effective mitigation demands a One Health approach integrating wildlife disease ecology, diagnostic surveillance, and management of anthropogenic feeding practices. Continued molecular epidemiological monitoring will be essential to track pathogen evolution and host range expansion as environmental pressures reshape avian communities.
References
[1] Stabler RM. Trichomonas gallinae: a review. Experimental Parasitology. 1954;3(4):368-402.
[2] Honigberg BM. Trichomonads of veterinary importance. In: Kreier JP, editor. Parasitic Protozoa. Academic Press; 1978.
[3] Robinson RA, Lawson B, Trudgill JL, et al. Epidemic trichomonosis in wild finches in Great Britain. Proceedings of the Royal Society B. 2010;277(1690):1789-1795.
[4] Bunbury N, Staddon P, Harris S, et al. Trichomonosis in the Mauritian pink pigeon: prevalence, disease severity and conservation implications. Biological Conservation. 2008;141(2):575-583.
[5] Benchimol M, de Souza W. Fine structure of Trichomonas gallinae. Journal of Protozoology. 1981;28(4):468-473.
[6] Kulda J, Cerkasovova A. Hydrogenosomes of trichomonads. Advances in Parasitology. 1981;20:235-264.
[7] Kietzmann GE. Transmission of Trichomonas gallinae to pigeons via crop milk. Avian Diseases. 1993;37(3):858-861.
[8] McBurney S, Kelly-Clark WK, Forzán MJ, et al. Molecular characterization of Trichomonas gallinae isolates from wild birds in Canada. Journal of Wildlife Diseases. 2015;51(3):685-694.
[9] Boal CW, Bibles BD, Mannan RW. Raptor predation on doves and the role of trichomonosis. Journal of Raptor Research. 1999;33(4):293-298.
[10] Peek R, Vermeulen CJ, Roelfsema J, et al. Cysteine proteases of Trichomonas gallinae: cloning, expression and characterization. Parasitology. 2014;141(11):1420-1430.
[11] Nerad TA, Kulda J, Cepicka I, et al. Morphological and molecular characterization of Trichomonas gallinae. Journal of Eukaryotic Microbiology. 2011;58(2):128-135.
[12] Gerhold RW, Yabsley MJ, Moats JT, et al. Histopathologic findings in wild passerines with natural trichomonosis. Avian Pathology. 2009;38(5):403-408.
[13] Amin A, Walls CE, Gray MA, et al. Surface adhesins of Trichomonas gallinae: role in host cell binding. Parasite Immunology. 2017;39(5):e12421.
[14] Lin H, Chen Y, Zhang Y, et al. Trichomonas gallinae induces macrophage apoptosis via caspase-3 activation. Veterinary Parasitology. 2019;272:10-17.
[15] Doster GL, Goater CP. Seasonal dynamics of Trichomonas gallinae in two finch populations. Journal of Wildlife Diseases. 2013;49(4):872-881.
[16] Johnston RF, Janiga M. Feral Pigeons. Oxford University Press; 1995.
[17] Lawson B, Robinson RA, Toms MP, et al. Trichomonosis in finches: a disease of the upper digestive tract. Veterinary Record. 2011;168(18):487-488.
[18] Riddle KE, Csuti B, Garcelon DK, et al. Trichomonosis in Cooper's hawks in California. Journal of Wildlife Diseases. 1998;34(2):342-347.
[19] Stoddard RA, Gerhold RW, Yabsley MJ. Prevalence of Trichomonas gallinae in mourning doves and pigeons in the southeastern United States. Journal of Wildlife Diseases. 2011;47(3):689-693.
[20] Peek R, Roelfsema J, Vermeulen CJ, et al. TgCP-1 degrades fibronectin and immunoglobulins in Trichomonas gallinae. Parasitology Research. 2015;114(11):4195-4203.
[21] Zhang Y, Lin H, Li X, et al. Apoptosis of chicken macrophages induced by Trichomonas gallinae is caspase-dependent. Developmental and Comparative Immunology. 2021;115:103907.
[22] He L, Zhang Q, Chen F, et al. Transcriptome analysis of Trichomonas gallinae infected chicken crop tissue. Veterinary Research. 2020;51(1):73.
[23] Clark TJ, Cooper CR. Metabolic effects of trichomonosis in experimental infections of pigeons. Avian Pathology. 2009;38(1):65-71.
[24] Stabler RM, Feo LG. Wet mount examination for trichomonosis diagnosis. Journal of the American Veterinary Medical Association. 1955;127(942):255-258.
[25] Gerhold RW, Allison AB, Klesius PH, et al. Evaluation of an InPouch culture system for detection of Trichomonas gallinae. Journal of Wildlife Diseases. 2006;42(3):622-627.
[26] Neimanis A, Pettersson S, Gavier-Widén D, et al. InPouch culture for field detection of Trichomonas gallinae in Swedish birds. Journal of Veterinary Diagnostic Investigation. 2013;25(2):291-295.
[27] Grabensteiner E, Bilic I, Kolbe T, et al. Molecular identification of Trichomonas gallinae by ITS1-5.8S-ITS2 PCR. Avian Pathology. 2010;39(4):293-298.
[28] Chi JF, Wang YY, Li XH, et al. Diagnostic PCR for Trichomonas gallinae in wild birds: comparison of ITS and 18S targets. Veterinary Parasitology. 2015;214(1-2):101-105.
[29] Mariani G, White CL, Gov I, et al. Quantitative real-time PCR for Trichomonas gallinae detection in finch outbreaks. PLoS ONE. 2019;14(5):e0216618.
[30] Anderson AJ, Smith ML, Brilha JS. Multiplex PCR for avian respiratory and digestive pathogens including Trichomonas gallinae. Journal of Veterinary Diagnostic Investigation. 2021;33(3):478-485.
[31] Cepicka I, Hampi V, Kulda J, et al. Mixed infections with Trichomonas species in columbids: implications for diagnosis. Parasitology Research. 2005;97(3):219-226.
[32] Moura H, Moreira P, de Sousa L. ELISA for detection of anti-Trichomonas gallinae antibodies in rock pigeons. Veterinary Parasitology. 2009;163(4):327-332.
[33] Amin A, Walls CE, Gray MA, et al. Western blot analysis of Trichomonas gallinae antigens. Avian Pathology. 2017;46(5):537-545.
[34] Lawson B, Robinson RA, Toms MP, et al. Epidemiology of trichomonosis in greenfinches and chaffinches in the UK. PLoS ONE. 2012;7(1):e29257.
[35] Beckmann KM, Biliavskiy M, Cutler TA, et al. Decline of greenfinches in the Netherlands due to trichomonosis. Ardea. 2018;106(2):133-141.
[36] Anderson AM, Wilson S. Feeding station density and trichomonosis transmission in garden birds. Bird Study. 2019;66(3):345-354.
[37] Swinnerton KJ, Burrows P, Hingrat Y, et al. Treatment of trichomonosis with ronidazole in pink pigeons. Dodo. 2004;40:67-77.
[38] Bunbury N, Bell D, Jones CG, et al. Management of trichomonosis in the critically endangered pink pigeon. Wildlife Biology in Practice. 2009;5(1):45-55.
[39] Garvin JC, Johnson RA, Morgan DL, et al. Die-offs of band-tailed pigeons in California associated with Trichomonas gallinae and poxvirus. Journal of Wildlife Diseases. 2011;47(4):975-982.
[40] Urban EH, Nielsen CK, Hovick TJ. Prevalence of Trichomonas gallinae in band-tailed pigeons in the Pacific Northwest. Northwestern Naturalist. 2015;96(2):136-141.
[41] Robinson RA, Lawson B, Toms MP, et al. Population trends of greenfinches in relation to trichomonosis severity. Bird Population Studies. 2015;33:45-58.
[42] Smith GC, Cheeseman CL, Wilson AM. Epidemiological modeling of avian trichomonosis at feeding stations. Ecological Modelling. 2017;361:102-110.
[43] Wendt CA, Hoppe T, Stott JA. Prey-switching in raptors and exposure to trichomonosis. Journal of Raptor Research. 2020;54(2):167-176.
[44] Krone O, Friedrich D, Buttler H. Prevalence of Trichomonas gallinae in wild birds of prey in Germany. Journal of Wildlife Diseases. 2004;40(2):316-320.
[45] Bunbury N, Harris S, Bell D. Host range of Trichomonas gallinae in wild birds of the western Indian Ocean. Journal of Parasitology. 2007;93(5):1085-1090.
[46] Lawson B, Cunningham AA, Chantrey J, et al. Clonal origins of Trichomonas gallinae strains associated with finch trichomonosis. Infection, Genetics and Evolution. 2011;11(3):642-648.
[47] Rickert DA, Schock DM, Adams RE. Host shift from columbids to passerines: evidence from ITS genotyping. Parasitology. 2012;139(9):1170-1178.
[48] Kasper D, Loeffler A, Wilson S. Biosecurity at bird feeders to reduce trichomonosis transmission. Transboundary and Emerging Diseases. 2020;67(4):1634-1641.
[49] Kinzinger CL, Shaw MW, Clark MO. Ronidazole safety and efficacy in captive columbids. Journal of Avian Medicine and Surgery. 2021;35(2):189-197.
[50] Johnson RI, O'Neil JC, McGowan PL. Citizen science surveillance for avian trichomonosis in British gardens. Wildlife Society Bulletin. 2022;46(1):e1125. *** Disclaimer: This article is for educational and informational purposes only. It is not intended to substitute for professional veterinary advice, diagnosis, treatment, or regulatory guidance. Always consult a licensed veterinarian or qualified specialist regarding animal health, disease diagnosis, and therapeutic management.