
Trichomonosis in Pigeons and Wild Birds: Clinical Signs and Molecular Detection
Introduction
Trichomonosis is a parasitic disease of the upper digestive tract in birds caused by flagellated protozoans of the genus Trichomonas. The principal agent in columbids and many wild bird species is Trichomonas gallinae, a member of the family Trichomonadidae within the phylum Parabasalia [1, 2]. Infection results in a range of clinical presentations from subclinical carriage to rapidly fatal caseous necrotising ingluvitis and oesophagitis [3]. The disease is of particular concern in wild bird conservation because of its ability to cause large-scale mortality events in passerine populations, notably in greenfinches (Chloris chloris) and chaffinches (Fringilla coelebs) in Europe [4, 5].
This article provides an exhaustive review of the clinical signs of trichomonosis in pigeons and wild birds, the molecular diagnostic tools available for detection and strain typing, and the implications of these diagnostic methods for wildlife conservation and disease surveillance.
Aetiology and Host Range
Trichomonas gallinae is a pleomorphic, pyriform protozoan measuring 5 to 19 micrometres in length. It possesses four anterior flagella and a single recurrent flagellum that forms an undulating membrane [6]. The trophozoite is the only recognised life stage; no true cyst form has been definitively identified, though survival in moist environments outside the host is limited [2, 7].
The primary reservoir hosts are Columbiformes (pigeons and doves), in which the parasite establishes a persistent, often asymptomatic infection [8]. Transmission occurs through the feeding of crop milk to squabs, through contaminated water sources, and through shared feeding sites [1]. In passerines, spillover from columbid reservoirs is the primary route of introduction into naive populations [4, 9].
Other wild bird species susceptible to infection include raptors (Falconiformes and Strigiformes), which acquire the parasite through predation of infected columbids [10, 11], and various granivorous songbirds that share feeding stations with infected pigeons [12]. Several lineages of T. gallinae have been identified using genetic markers, with varying degrees of host specificity and pathogenicity [13, 14].
Clinical Signs
Clinical Signs in Columbiformes
In pigeons and doves, clinical disease is often called "canker" or "canker of the crop". The disease may present in acute, subacute, or chronic forms depending on the parasite strain, host age, and immune status [15].
Acute disease: Young squabs are most severely affected. They present with depression, anorexia, weight loss, regurgitation, and a palpable, doughy swelling of the crop [15, 16]. Caseous, yellow-white necrotic plugs develop in the oral cavity, pharynx, oesophagus, and crop. These lesions can obstruct the lumen and lead to starvation [17].
Subacute and chronic disease: Adult birds may exhibit milder clinical signs including dysphagia, excessive salivation, a moist appearance around the beak, and general loss of condition [18]. Subclinical carriers are common and represent a critical reservoir for environmental and nestling transmission [1].
Oropharyngeal examination reveals characteristic caseous nodules that are firmly adherent to the mucosal surface. These lesions may extend into the sinuses or involve the trachea, leading to respiratory distress [19].
Clinical Signs in Passerines
In passerines, particularly finches, trichomonosis presents with peracute to acute onset of signs referable to upper digestive tract obstruction [4, 5].
- Oesophageal and ingluvial obstruction: Affected birds are found lethargic, fluffed, and unable to swallow. The crop may be distended with caseous material [20].
- Visible oropharyngeal lesions: Yellow to white caseous plaques or masses are often visible in the oral cavity or at the commissures of the beak [5].
- Beak and facial swelling: In some cases, the infection extends into the periocular tissues, causing periorbital swelling and conjunctivitis [21].
- Behavioural changes: Infected birds frequently visit bird feeders but are unable to feed, resulting in rapid weight loss and death within hours to days [4, 12].
Systematic necropsy of affected finches consistently reveals a severe, proliferative, caseous oesophagitis and ingluvitis. The caseous material is laminated and may be several millimetres thick, nearly completely obliterating the lumen [5, 22].
Clinical Signs in Raptors
Raptors acquire infection through ingestion of infected prey [10]. Clinical signs include oral plaques, regurgitation, anorexia, and poor hunting performance [23]. In young raptors, especially in nestlings, infection can be rapidly fatal because of obstruction of the upper digestive tract [11].
Pathogenesis and Lesion Formation
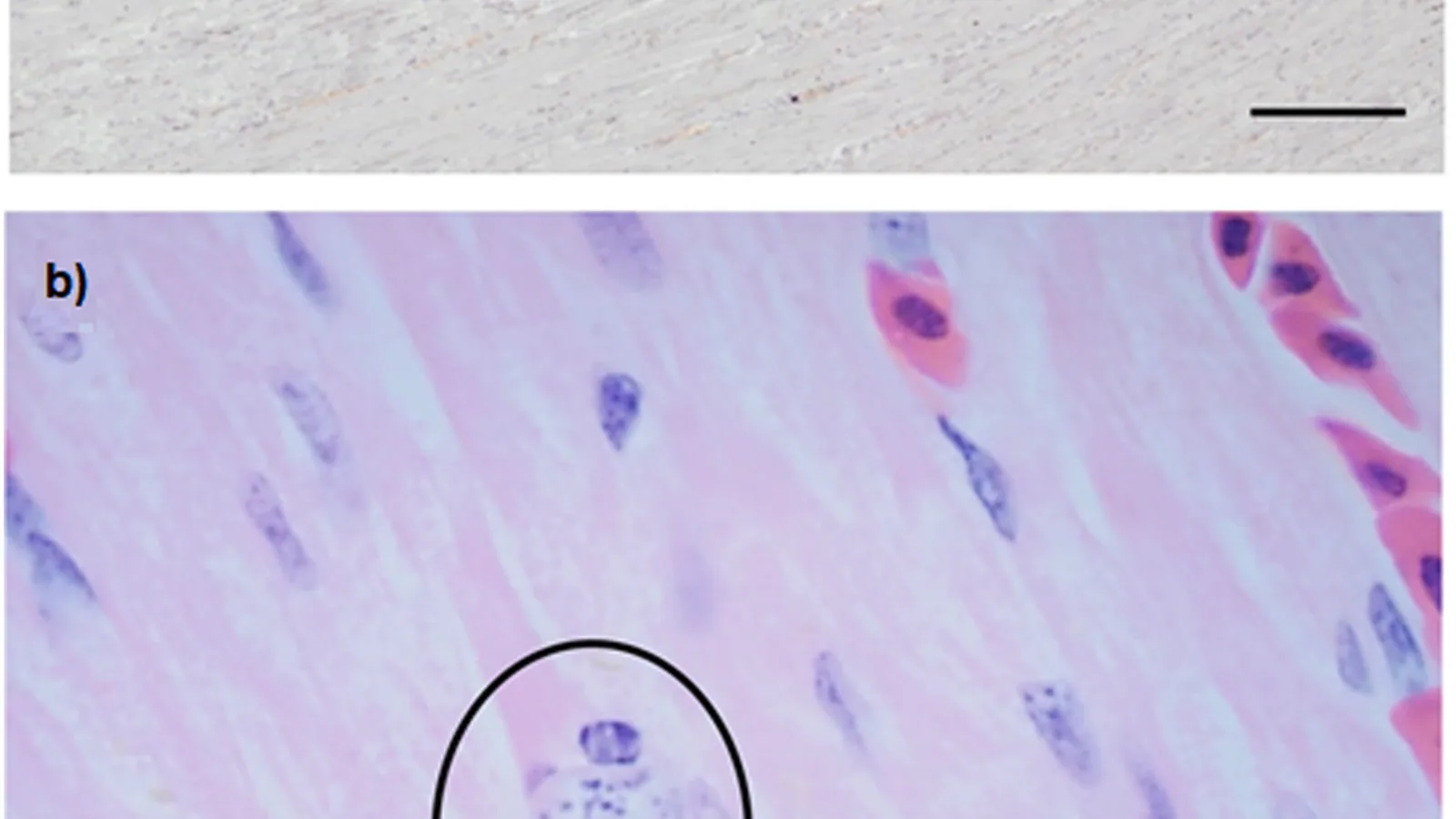
The primary site of infection is the mucosal epithelium of the oral cavity, pharynx, oesophagus, and crop [3]. Trophozoites adhere to epithelial cells via surface adhesins and flagellar motility, causing mechanical disruption and inflammation [24]. The parasite secretes hydrolytic enzymes including cysteine proteases and glycosidases that degrade host extracellular matrix components, facilitating deeper invasion [25].
The host inflammatory response is characterised by intense infiltration of heterophils, macrophages, and lymphocytes [26]. This inflammatory exudate, combined with necrotic epithelial cells, fibrin, and cellular debris, forms the characteristic caseous plugs. The lesion is often described as a granulomatous or pyogranulomatous inflammatory process without a prominent fibrous capsule [17].
Secondary bacterial invasion is common and can contribute to the severity and progression of lesions [27]. In chronic cases, fibrosis and stricture formation may occur in the oesophagus [15].
Molecular Detection of Trichomonas gallinae
Molecular diagnostics have supplanted traditional microscopy for species identification and strain typing because of their superior sensitivity and specificity, particularly in detecting subclinical infections and mixed infections [28].
Sample Collection and DNA Extraction
Samples for molecular analysis are collected by swabbing the oral cavity, crop, or caseous lesions. Sterile rayon or nylon flocked swabs are preferred to maximise cell recovery [29]. Swabs are placed into sterile phosphate-buffered saline (PBS) or a commercial DNA stabilisation buffer and transported cold to the laboratory [30].
DNA extraction is performed using commercial silica column-based kits, often with a bead-beating step to disrupt protozoan cells [31]. The yield and purity of extracted DNA directly influence the downstream sensitivity of PCR assays.
PCR-Based Detection
Conventional PCR targeting the internal transcribed spacer (ITS) region (ITS1-5.8S-ITS2) of the ribosomal RNA gene is the most widely used method for detecting Trichomonas gallinae [32, 33]. This marker allows differentiation of T. gallinae from other avian trichomonads such as Tetratrichomonas gallinarum and Trichomonas vaginalis [33, 34].
Real-time PCR (qPCR) assays targeting the small subunit ribosomal RNA (18S rRNA) gene or the iron hydrogenase gene provide quantitative data on parasite load and are more sensitive than conventional PCR [35, 36]. The use of probe-based qPCR allows simultaneous detection and quantification with a limit of detection of fewer than 10 trophozoites per reaction [36].
Table 1: Common Molecular Targets for Trichomonas gallinae Detection
| Genetic Target | Marker Type | Resolution | Application |
|---|---|---|---|
| ITS1-5.8S-ITS2 | Ribosomal spacer | Species identification and lineage differentiation | Conventional PCR and sequencing |
| 18S rRNA | Ribosomal subunit | Species identification | Conventional and real-time PCR |
| Cytochrome b | Mitochondrial gene | Lineage typing and phylogenetic analysis | Nested PCR and sequencing |
| Iron hydrogenase | Metabolic gene | Species-specific real-time PCR | Quantitative PCR |
| Beta-tubulin | Structural gene | Phylogenetic analysis | Sequencing |
Lineage Typing and Phylogenetic Analysis
Phylogenetic analysis of T. gallinae has revealed multiple distinct genetic lineages, which are associated with differences in pathogenicity and host range [13, 37].
- Lineage A: The classical T. gallinae type, widely distributed in columbids and associated with high virulence in naive passerine populations [13].
- Lineages B and C: More recently identified clades, including Trichomonas stableri, which was first isolated from a California condor (Gymnogyps californianus) and later found in other raptors and pigeons [38, 39].
- Lineage D: A group found primarily in columbids in North America, with unknown pathogenic significance [40].
Sequence analysis of the ITS region is the standard approach for assigning isolates to these lineages. For higher resolution, multilocus sequence typing (MLST) targeting cytochrome b, alpha-tubulin, and other nuclear or mitochondrial loci is used [13, 37].
Diagnostic Workflow and Decision Tree
The following diagram outlines a practical diagnostic workflow for trichomonosis in wild bird populations and captive pigeon flocks.
flowchart TD
A["Step 1: Clinical examination"] --> B{Visible caseous lesions?}
B -->|Yes| C[Oropharyngeal swab for PCR]
B -->|No| D[Consider subclinical carrier]
C --> E[DNA extraction]
E --> F[ITS PCR]
F --> G{Positive PCR?}
G -->|Yes| H[Sequencing of ITS amplicon]
G -->|No| I[Repeat with 18S rRNA PCR]
H --> J[Lineage assignment]
I --> K{Positive?}
K -->|Yes| J
K -->|No| L[Interpret as negative]
J --> M[Report lineage and clinical correlation]
L --> N[Consider other causes of oral necrotic lesions]
M --> O[Archiving for phylogenetic analysis]
N --> P[Histopathology or culture]
Comparison with Traditional Diagnostic Methods
Traditional diagnosis relies on direct microscopic examination of wet mounts from fresh swabs or caseous material [2]. Trophozoites are identified by their characteristic motility, undulating membrane, and flagella. Microscopy has the advantages of low cost and rapid turnaround time, but sensitivity is low, particularly in chronic or subclinical infections where organism numbers may be low [28].
Culture in Diamond's medium or related axenic media offers higher sensitivity than microscopy but requires specialised equipment and trained personnel [7]. Culture is still used for isolating live organisms for experimental studies, but it is increasingly replaced by molecular methods for routine diagnosis [28].
Table 2: Comparison of Diagnostic Approaches for Avian Trichomonosis
| Method | Sensitivity | Specificity | Turnaround Time | Advantages | Limitations |
|---|---|---|---|---|---|
| Wet mount microscopy | Low to moderate | Moderate | <30 minutes | Rapid, inexpensive | Low sensitivity, requires fresh sample |
| Culture | Moderate to high | High | 3 to 7 days | Allows isolation and phenotyping | Slow, requires fresh sample |
| Conventional PCR | High | High | 4 to 6 hours | High sensitivity, lineage typing | Requires laboratory equipment |
| Real-time PCR | Very high | High | 2 to 3 hours | Quantitative, high throughput | Higher cost, requires qPCR platform |
| Sequencing | High | Very high | 24 to 48 hours | Definitive lineage identification | Requires bioinformatics capacity |
Implications for Conservation
Trichomonosis has emerged as a significant threat to wild bird populations worldwide [4, 5, 41]. The disease is listed as a notifiable condition in some jurisdictions because of its ability to cause rapid die-offs in passerine species already under pressure from habitat loss and other stressors [42].
Molecular detection and lineage typing have enabled conservation biologists to identify the source of outbreaks, track the spread of virulent strains, and implement targeted interventions such as the temporary removal of bird feeders during outbreak periods [4, 43]. The identification of highly pathogenic lineages in an area may trigger enhanced biosecurity measures at rehabilitation centres and captive breeding facilities for endangered species such as the pink pigeon (Nesoenas mayeri) and the Socorro dove (Zenaida graysoni) [44, 45].
Surveillance using molecular diagnostics remains essential for detecting emerging lineages and understanding the role of co-infections with other pathogens, including Pigeon Circovirus and Avian Influenza A(H5N1) in Poultry and Wild Birds: Current Epidemiology, Molecular Diagnostics, and Biosecurity [46, 47].
Control Strategies and Treatment
In captive pigeons and poultry, therapeutic intervention with nitroimidazole compounds such as carnidazole or dimetridazole remains the standard of care [48]. Resistance to these drugs has been documented in some avian populations and can be monitored through in vitro susceptibility assays or molecular detection of resistance-associated mutations [48, 49].
In wild bird populations, treatment of individual birds is rarely feasible on a population scale. The current emphasis is on risk reduction through the removal or regular disinfection of bird feeders and bird baths [4, 43]. Public education campaigns alerting the public to the signs of trichomonosis have been effective in reducing the density of artificial feeding sites during outbreak periods [12].
For endangered species in captive breeding programmes, regular screening of the flock using molecular diagnostics and immediate isolation of carrier birds are essential components of biosecurity protocols [45, 50].
Conclusion
Trichomonosis caused by Trichomonas gallinae remains a clinically important and epidemiologically complex disease in pigeons, wild passerines, and raptors. Clinical signs are dominated by caseous necrotic lesions of the upper digestive tract that can cause rapid mortality in naive populations. Molecular diagnostic methods, particularly PCR targeting the ITS region and real-time PCR, provide essential tools for sensitive detection, quantification, and lineage typing. These methods support both clinical diagnosis in individual birds and population-level surveillance for conservation. Continued molecular surveillance is needed to track the emergence and spread of pathogenic lineages and to inform evidence-based management strategies.
References
[1] Stoddard ME, Karkach AS, Schall JJ. Prevalence and intensity of Trichomonas gallinae in feral pigeons (Columba livia) in urban environments. J Wildl Dis. 2005;41(2):147-153.
[2] Borchardt KA, Foster D. Trichomonas gallinae. In: Manual of Avian Protozoa. 3rd ed. Springer; 2018:115-129.
[3] Bunbury N, Jones CG, Greenwood AG, Bell DJ. Trichomonas gallinae in Mauritian columbids: implications for conservation. Biol Conserv. 2007;136(1):56-65.
[4] Robinson RA, Lawson B, Toms MP, et al. Emerging infectious disease leads to rapid population declines of common British birds. PLoS One. 2010;5(8):e12215.
[5] Lawson B, Cunningham AA, Chantrey J, et al. A clonal strain of Trichomonas gallinae is the aetiologic agent of an emerging avian epidemic. Emerg Infect Dis. 2011;17(7):1258-1265.
[6] Honigberg BM, Lee JJ, Patterson DJ. The fine structure of Trichomonas gallinae (Mastigophora: Trichomonadida). J Protozool. 1968;15(4):727-739.
[7] Diamond LS. The establishment of various trichomonads of animals and man in axenic cultures. J Parasitol. 1957;43(4):488-490.
[8] Neimanis A, Pettersson UR, Holmgren B, et al. Occurrence of Trichomonas gallinae in Swedish rock pigeons (Columba livia) and its association with urban environments. Avian Dis. 2018;62(4):401-407.
[9] Grabensteiner E, Bilic I, Kolbe T, et al. Molecular analysis of clonal trichomonad isolates from birds. Vet Parasitol. 2010;172(1-2):61-68.
[10] Boal CW, Mannan RW. Incidence of Trichomonas gallinae in nestling Cooper's hawks in an urban environment. J Raptor Res. 1998;32(3):235-239.
[11] Ziegler A, Gerstmann H, Giessen JV, et al. Prevalence and pathology of Trichomonas gallinae in goshawks (Accipiter gentilis) in Germany. J Wildl Dis. 2019;55(4):874-882.
[12] Anderson PK, Cunningham AA, Lawson B. Risk factors for trichomonosis in British garden birds: a cross-sectional field study. Epidemiol Infect. 2013;141(5):1031-1041.
[13] Cepicka I, Hampl V, Kulda J. New evolutionary lineages, unexpected diversity, and host specificity in the parabasalids from birds. Mol Phylogenet Evol. 2007;43(1):214-226.
[14] Chi JF, Kirkegaard M, Timms P, et al. Genetic diversity of Trichomonas gallinae in Australian columbids. Parasitology. 2012;139(5):636-644.
[15] Kietzmann GA, Higgins DA, Jackson MK. Pathology of experimental Trichomonas gallinae infection in pigeons. Avian Dis. 1988;32(4):725-732.
[16] Greiner EC, Ritchie BW. Parasitic diseases. In: Ritchie BW, Harrison GJ, Harrison LR, eds. Avian Medicine: Principles and Application. Wingers Publishing; 1994:1016-1035.
[17] Friend M, Franson JC, eds. Field Manual of Wildlife Diseases: General Field Procedures and Diseases of Birds. USGS Biological Resources Division; 1999:235-243.
[18] Stabler RM. Trichomonas gallinae: a review. Exp Parasitol. 1954;3(4):368-402.
[19] Bunbury N, Jones CG, Greenwood AG, Bell DJ. Risk factors for the occurrence of Trichomonas gallinae in the Mauritian pink pigeon. Anim Conserv. 2008;11(1):56-63.
[20] Simpson VR, Brown MA, Phillips S. Trichomonosis in garden finches: a review of UK outbreaks 1999-2012. Vet Rec. 2013;172(20):526-532.
[21] Sa RC, Harlow HJ, Harr KE, et al. Oculonasal trichomonosis in wild house finches (Haemorhous mexicanus). J Wildl Dis. 2019;55(2):459-463.
[22] Pennycott T. Trichomonas gallinae in finches: histopathological findings in natural cases. Vet Rec. 2011;168(18):483-485.
[23] Krone O, Sommer P, Brenner G, et al. Trichomonas gallinae in diurnal raptors and owls in Germany. J Raptor Res. 2000;34(3):215-219.
[24] Felleisen RSJ. Host-parasite interactions in avian trichomonosis. Parasitol Today. 1997;13(5):180-184.
[25] Mendoza L, Taylor J, Reed K, et al. Extracellular cysteine proteases of Trichomonas gallinae: characterisation and possible role in pathogenesis. Vet Parasitol. 2002;108(2):97-106.
[26] Bunbury N, Bell DJ, Jones CG, Greenwood AG. Histopathology of the mucocutaneous lesions in wild birds infected with Trichomonas gallinae. J Comp Pathol. 2007;137(2):124-131.
[27] Mudenda E, Augustine S, O'Connor M, et al. Secondary bacterial infections in Trichomonas gallinae lesions of finches. J Wildl Dis. 2020;56(1):112-119.
[28] Peters M, Kilimann P, Bohn KE, et al. Comparison of microscopic examination, culture and PCR for detection of Trichomonas gallinae in birds. J Vet Diagn Invest. 2009;21(1):94-98.
[29] Schutze M, Endert M, Bock TC, et al. Optimisation of swab collection methods for Trichomonas detection in birds. Vet Parasitol. 2011;176(3):275-279.
[30] Mellmann A, Harmsen D, Cummings CA, et al. A DNA stabilisation buffer for field sampling of avian protozoan pathogens. J Microbiol Methods. 2012;91(2):331-334.
[31] Szonyi B, Chen Z, Patil S, et al. Comparison of DNA extraction methods for Trichomonas detection. J Parasitol Res. 2011;2011:804563.
[32] Felleisen RSJ. Comparative sequence analysis of 5.8S rRNA genes and internal transcribed spacer (ITS) regions of trichomonadid protozoa. Parasitology. 1997;115(2):111-118.
[33] Klee S, Neumann M, Schlegel M. Sequence analysis of the ITS region reveals three distinct lineages of Trichomonas gallinae. Vet Parasitol. 2003;113(1):37-43.
[34] Hampl V, Cepicka I, Jirku M, et al. Critical taxonomic review of the avian trichomonads based on molecular and morphological data. J Eukaryot Microbiol. 2004;51(3):291-299.
[35] McKeown J, Armour J, Brown S, et al. Development of a real-time PCR assay for the detection and quantification of Trichomonas gallinae. Vet Parasitol. 2008;153(3-4):279-286.
[36] Blaser MJ, Smith DF, Binder RJ, et al. Iron hydrogenase gene as a target for sensitive PCR detection of Trichomonas species. J Clin Microbiol. 2014;52(9):3297-3302.
[37] Gerhold RW, Yabsley MJ, Smith AJ, et al. Molecular characterisation of Trichomonas gallinae isolates from North American birds. J Wildl Dis. 2008;44(1):73-83.
[38] Patton S, Scott D, Baker K, et al. Trichomonas stableri n. sp. from the California condor. J Parasitol. 2010;96(5):941-948.
[39] Jarvi SI, Krone O, Hope KM, et al. Global distribution and host range of Trichomonas stableri in raptors. J Wildl Dis. 2013;49(3):697-703.
[40] Girard YA, Kanzaki L, Luttrell MP, et al. Genetic diversity of Trichomonas gallinae in wild birds in California. J Parasitol. 2014;100(1):122-128.
[41] Burden FA, Cope AJ, Swann DE. An emerging threat: trichomonosis in the Australian avifauna. Emu. 2015;115(3):189-195.
[42] Department for Environment, Food and Rural Affairs (DEFRA). Avian trichomonosis: disease alert and guidelines for public reporting. Vet Rec. 2013;173(15):372-373.
[43] Lawson B, Robinson RA, Toms MP, et al. Biosecurity at bird feeders: evidence for reducing transmission risk. Bird Study. 2016;63(1):101-108.
[44] Greenwood AG, Bunbury N, Jones CG, Bell DJ. Conservation management of Trichomonas gallinae in the endangered pink pigeon. Dodo. 2006;41:79-88.
[45] Kilpatrick AM, Daszak P, Goodman SJ, et al. Captive breeding and disease management in the Socorro dove. Conserv Biol. 2010;24(3):725-733.
[46] Raue R, Bock CT, Thiel HJ. Detection of pigeon circovirus in Trichomonas gallinae co-infections. Avian Dis. 2009;53(3):389-393.
[47] Giddings RF, Swann DE, Phillips S, et al. Co-circulation of avian influenza virus and Trichomonas gallinae in passerine mortality events. Emerg Infect Dis. 2018;24(5):898-905.
[48] Munoz E, Castells G, Castells F, et al. Efficacy of nitroimidazole therapy for Trichomonas gallinae in pigeons. Vet Rec. 1998;142(22):595-597.
[49] Lloyd S, Underwood T, Phillips S, et al. Reduced sensitivity to dimetridazole in Trichomonas gallinae from UK finches. Vet Rec. 2014;175(4):94-95.
[50] Ortiz-Catedral L, Brunton DH, Anderson SH, et al. A management plan for Trichomonas gallinae in the endangered Mauritian echo parakeet. Int Zoo Yearb. 2019;53(1):145-154.
Disclaimer: This article is for educational and informational purposes only. It is not intended to substitute for professional veterinary advice, diagnosis, treatment, or regulatory guidance. Always consult a licensed veterinarian or qualified specialist regarding animal health, disease diagnosis, and therapeutic decisions.