
Infectious Salmon Anaemia Virus (ISAV)
Introduction and Classification
Infectious salmon anaemia virus (ISAV) is the causative agent of infectious salmon anaemia (ISA), a notifiable disease of farmed Atlantic salmon (Salmo salar) [1]. ISAV is classified within the family Orthomyxoviridae and is the type species of the genus Isavirus [1]. The virus was first identified in Norway in the mid-1980s and has since been reported in Canada, Chile, Scotland, the Faroe Islands, and other salmon-producing regions [2, 1]. Unlike other orthomyxoviruses such as influenza A virus, ISAV is restricted to aquatic hosts and primarily infects teleost fish [1].
The ISAV genome consists of eight segments of single-stranded negative-sense RNA, totaling approximately 14.5 kb [3, 1]. Each segment is flanked by non-coding regions (NCRs) that contain regulatory elements for transcription and replication [3]. Bioinformatic analyses of the NCRs have identified conserved nucleotide positions and secondary structures likely involved in binding by the RNA-dependent RNA polymerase (RdRp) and ribosomal recognition of mRNA [3]. Two major genetic groups exist: European (EU) and North American (NA), based on phylogenetic analyses of the hemagglutinin-esterase (HE) gene and other segments [2, 32].
Genomic Organization and Viral Proteins
The eight genomic segments encode at least ten proteins [4, 1]. Segment 1 encodes the PB2 polymerase subunit (RdRp component). Segment 2 encodes PB1. Segment 3 encodes PA. Segment 4 encodes the nucleoprotein (NP). Segment 5 encodes the fusion protein (F), which is cleaved by host proteases and is a key determinant of virulence [1, 5]. Segment 6 encodes the hemagglutinin-esterase (HE) protein, which mediates receptor binding and receptor-destroying activity [1]. Segment 7 encodes two proteins via alternative splicing: the matrix protein (M) and a nuclear export protein (NEP) [4]. Segment 8 encodes the non-structural protein NS1 and the nuclear export protein (NS2/NEP) [1].
The highly polymorphic region (HPR) of the HE gene (segment 6) is central to ISAV virulence [1, 31]. Viruses with a full-length HPR (designated HPR0) are considered non-virulent, while those with deletions in the HPR (designated HPRΔ) are virulent [1, 33]. The transition from HPR0 to HPRΔ involves intra-segmental recombination via template switching, leading to sequence deletions and insertions [31]. Additional mutations in the fusion protein (segment 5), such as a Q266L substitution or microdeletion in the putative protease cleavage site, are also required for full virulence [1, 5].
| Genomic Segment | Encoded Protein(s) | Function | Key References |
|---|---|---|---|
| 1 | PB2 | RNA-dependent RNA polymerase subunit | [1] |
| 2 | PB1 | RNA-dependent RNA polymerase subunit | [1] |
| 3 | PA | RNA-dependent RNA polymerase subunit | [1] |
| 4 | Nucleoprotein (NP) | RNA encapsidation | [1] |
| 5 | Fusion protein (F) | Membrane fusion, virulence determinant | [1, 5, 31] |
| 6 | Hemagglutinin-esterase (HE) | Receptor binding, receptor-destroying, HPR | [1, 31] |
| 7 | Matrix protein (M), Nuclear export protein (NEP) | Virion structure, nuclear export of vRNPs | [4, 1] |
| 8 | NS1, NS2/NEP | Interferon antagonism, nuclear export? | [1] |
Virion Structure and Morphology
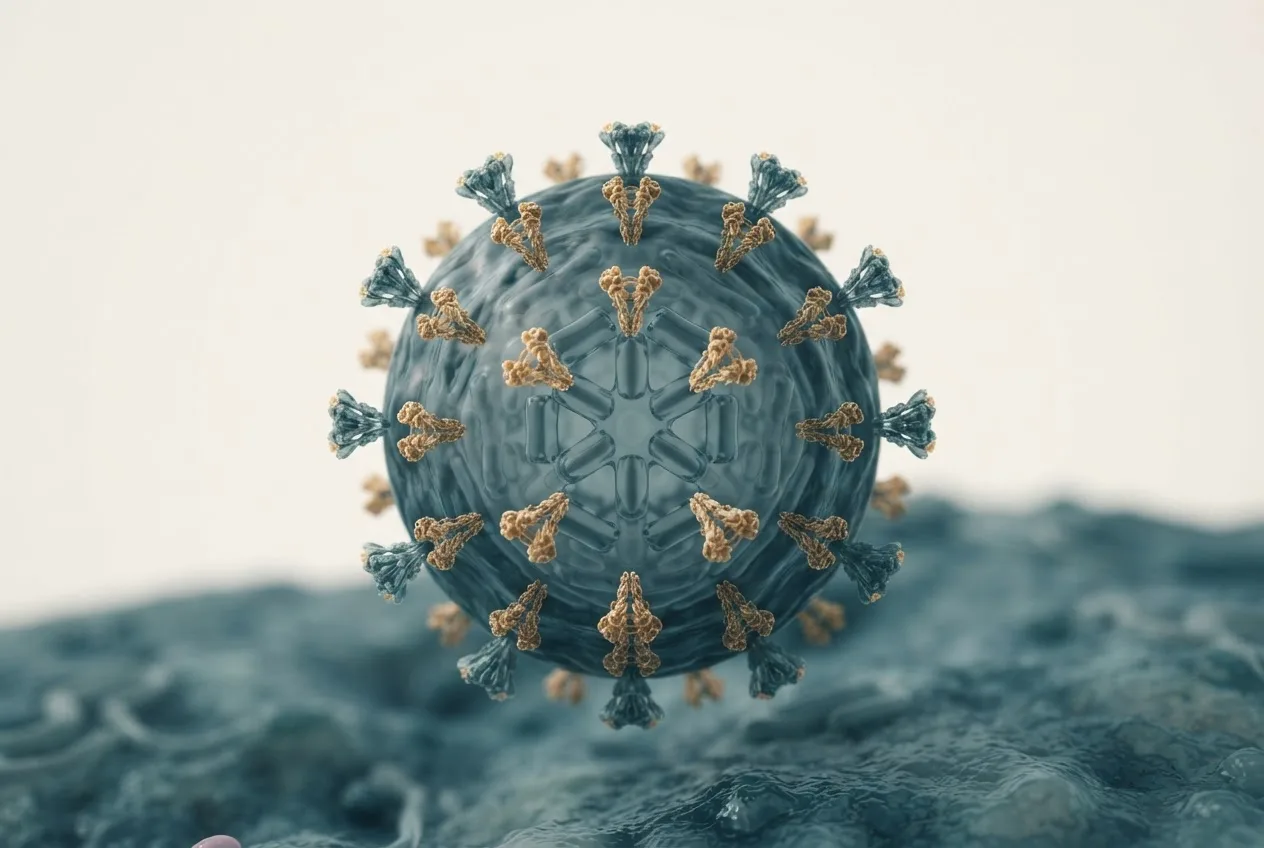
ISAV virions are enveloped, pleomorphic, and range from 80 to 140 nm in diameter [6]. The envelope is derived from the host cell plasma membrane and contains two integral membrane glycoproteins: HE and F [1]. The virion surface displays 10–12 nm spikes composed of HE and F trimers [6]. Internally, the viral genome is encapsidated by NP into helical ribonucleoprotein complexes (vRNPs) associated with the RdRp complex (PB1, PB2, PA) [1]. The matrix protein (M) lines the inner leaflet of the envelope and provides structural integrity [4].
Morphogenesis occurs at the plasma membrane of infected endothelial cells [6]. Virions bud from the cell surface and are released into the extracellular space. Electron microscopy studies have shown filamentous and spherical forms of ISAV particles [6]. The fusion protein F is activated by proteolytic cleavage at a monobasic or multibasic site, depending on the isolate, which confers the ability to fuse with host cell endosomal membranes at low pH [1, 7].
Viral Entry and Replication Cycle
ISAV entry into host cells occurs via endocytosis, involving both clathrin-mediated endocytosis and macropinocytosis [8]. Pharmacological inhibitors of clathrin-mediated uptake (e.g., chlorpromazine) and inhibitors of Na+/H+ exchange (e.g., amiloride, which affects macropinocytosis) significantly reduce infection in Atlantic salmon head kidney (ASK) cells [8]. Virus particles are observed in coated pits and membrane ruffles, the latter induced by actin rearrangement [8]. After internalization, low pH within the endosome triggers conformational changes in the F protein, exposing the fusion peptide and mediating fusion between the viral envelope and the endosomal membrane [7]. The fusion peptides of ISAV adopt a β-sheet secondary structure and are capable of fusing lipid vesicles in the presence or absence of cholesterol, though cholesterol modulates fusion efficiency [7].
Following fusion, vRNPs are released into the cytoplasm and imported into the nucleus, where transcription and replication occur [1]. The viral RdRp synthesizes capped mRNAs using cap-snatching from host pre-mRNAs (as in influenza viruses [3]). The NEP protein (encoded by segment 7) mediates nuclear export of progeny vRNPs [4]. Replication of the genome proceeds via a complementary RNA (cRNA) intermediate [3]. Progeny vRNPs are transported to the plasma membrane, where assembly and budding occur [6, 1].
Pathogenesis and Target Cells
The primary target cells of virulent ISAV (HPRΔ) are endothelial cells lining blood vessels and heart endocardium [9, 28]. Infection causes widespread endothelial damage, leading to hemorrhages, edema, and exophthalmia [10, 11]. The virus also infects mononuclear phagocytes, as demonstrated by single-nucleus RNA sequencing [9]. In Atlantic salmon, ISAV infection results in severe anaemia, which is a hallmark of the disease [10, 12]. Erythrocytes bind ISAV via the HE protein and sequester infective virus particles, but do not support active replication in most cases [12]. However, a small fraction of erythrocytes may express viral proteins, suggesting occasional internalization and transcription [12].
Ocular manifestations of ISAV infection include corneal dysplasia, edema, and changes in the choroid rete mirabile [10]. Immunohistochemistry reveals ISAV antigen accumulation in corneal stroma, and dysregulation of IgM, Sox6, Sox9, collagen type 1, Gata1, and CD10 expression in ocular tissues [10]. The anaemia is reflected in decreased erythrocyte complement within the choroid rete, accompanied by compensatory upregulation of Gata1, a key erythropoiesis regulator [10].
In contrast, the non-virulent HPR0 variant shows epithelial tropism, targeting gill epithelial cells rather than endothelial cells [28]. HPR0 infection is localized and transient, typically not causing systemic disease [25, 28].
Comparative studies in rainbow trout (Oncorhynchus mykiss) show that ISAV induces similar but less severe lesions than in Atlantic salmon, including endothelial necrosis and haemorrhages [11].
Host Range and Susceptibility
The primary susceptible species is Atlantic salmon (Salmo salar), but other species can be infected [13]. Rainbow trout (Oncorhynchus mykiss) are susceptible to experimental infection and show pathological changes, though mortality is variable [11, 35]. Coho salmon (Oncorhynchus kisutch) have been identified as hosts in Chile [14]. Sea trout (Salmo trutta) can act as carriers after experimental infection and may shed the virus [15]. Herring (Clupea harengus) have also been shown to harbor ISAV, suggesting a potential wild reservoir [13].
Lumpfish (Cyclopterus lumpus), commonly used as cleaner fish in salmon aquaculture, do not appear to be susceptible to ISAV [16]. Cohabitation and intraperitoneal challenges failed to produce systemic infection or disease, although occasional viral RNA was detected in gills and blood, likely due to environmental contamination [16].
Wild Atlantic salmon returning to Norwegian rivers have been found to carry ISAV-HPR0 at low prevalence, and phylogenetic analyses indicate that wild salmon may serve as a reservoir for low-virulent variants [32].
Epidemiology and Transmission
ISAV is transmitted horizontally via water and direct contact [17, 18]. The virus can survive and remain infectious in seawater for extended periods, with survival kinetics influenced by temperature, salinity, and organic material [18]. Environmental surveillance using charged membrane filters and buffers has been developed to concentrate and detect ISAV from seawater samples [26]. The estimated limit of quantification is 2.2 x 10³ ISAV copies per liter of natural seawater [26].
Transmission also occurs via contaminated equipment, personnel, and possibly through sea lice (Lepeophtheirus salmonis) [30]. Vertical transmission via fertilized eggs and gametes is considered unlikely, though surface contamination cannot be excluded [19, 25]. EFSA risk assessments indicate that disinfection of fertilised eggs reduces the probability of introduction of HPRΔ ISAV into free areas to near zero [19].
Outbreaks typically occur in marine farm sites with high stocking densities [2, 17]. Descriptive epidemiology from Newfoundland revealed concurrent detection of multiple ISAV variants (both HPR0 and HPRΔ) on the same farm, with mortality increases following rises in viral prevalence [2]. Both European and North American variants can co-circulate on a single farm [2].
The non-virulent HPR0 variant is widespread in salmon aquaculture and is considered the progenitor of virulent HPRΔ variants [1, 33]. The transition from HPR0 to HPRΔ has been observed in field outbreaks, likely triggered by deletions in the HPR region via recombination [31, 33]. Low-virulent variants can persist in farmed populations and serve as a continuous source of new virulent strains [30].
Diagnosis
Diagnosis of ISAV infection relies on molecular detection of viral RNA, histopathology, immunohistochemistry, and virus isolation [15, 20]. Real-time reverse transcription polymerase chain reaction (RT-qPCR) targeting the HE gene is the gold standard for screening [2, 15, 20]. A Bayesian analysis of diagnostic sensitivity and specificity for ISAV detection in Atlantic Canada compared IFAT (indirect fluorescent antibody test) and real-time RT-PCR, with both methods showing high sensitivity and specificity [20].
Sequencing of segment 6 and segment 5 is performed to differentiate HPR0 from HPRΔ variants and to identify virulence markers [2, 1, 32]. Genotyping of the HPR region allows assignment to European or North American clades [2, 32].
Virus isolation is possible in cell lines such as SHK-1 (salmon head kidney) and ASK (Atlantic salmon kidney) [21, 8, 22]. Historically, HPR0 variants resisted propagation in cell culture, but a successful isolation of an atypical HPR0 variant with a virulence marker in segment 5 has been reported, challenging previous assumptions [27].
Immunohistochemistry and immunofluorescence can detect viral antigen in tissues, especially in gill epithelial cells for HPR0 and endothelial cells for HPRΔ [28].
Genetic Diversity and Evolution
ISAV evolves through point mutations, segment reassortment, and recombination [1, 31]. Reassortment between strains of different genetic groups (e.g., European and North American) can generate novel viruses with altered virulence [2]. Intra-segmental recombination, particularly in the HPR, is driven by template switching during RNA replication and can create both deletions and insertions [31]. Experimental studies using recombinant ISAV (rISAV) with different HPR genotypes confirmed rapid sequence modifications in the HPR, with inserted sequences originating from segments 1, 5, 6, and 8 [31].
Phylogenetic analyses of the HE gene classify ISAV into four major clades: three European (EU-1 to EU-3) and one European-like from northeastern North America (EU-NA), plus divergent North American lineages [32]. Virulent HPRΔ variants appear across these clades, supporting the hypothesis that HPR0-to-HPRΔ transitions occur frequently in farmed populations [30, 33].
Control and Prevention
Control of ISA relies on biosecurity measures, stamping out of infected populations, movement restrictions, and surveillance [2, 30]. Vaccination is practiced in some regions, but vaccines are only partially effective and do not prevent infection [1]. Selective breeding for genetic resistance is a promising approach, with heritability estimates of 0.13–0.33 for binary survival [29]. Genome-wide association studies have identified a quantitative trait locus on chromosome Ssa13 that explains 3% of genetic variance, but resistance is polygenic [29]. RNA sequencing of heart tissue from resistant and susceptible fish revealed upregulation of TRIM25 in resistant individuals [29].
Gene editing approaches, such as triple gene disruption, have been explored in experimental settings to confer genetic resistance [23]. However, such interventions are not yet commercially applied.
Monovalent vaccines based on inactivated virus or recombinant proteins are available but do not provide sterile immunity [1]. Management focuses on early detection through active surveillance and depopulation of infected cages.
Mermaid Diagram: ISAV Pathogenesis and Transmission Flowchart
flowchart TD
A[Infected Atlantic salmon], >|Horizontal transmission via water| B(Waterborne ISAV)
B, > C(Entry via gills<br/>or skin)
C, > D{Primary target}
D, >|HPR0| E[Gill epithelial cells]
D, >|HPRΔ| F[Endothelial cells<br/>(blood vessels, heart)]
E, > G(Localized infection,<br/>no systemic disease)
F, > H(Systemic endothelial infection)
H, > I(Endothelial damage, hemorrhages)
I, > J[Anaemia, exophthalmia, corneal lesions]
J, > K(Mortality)
H, > L(Virus shedding via feces, urine, mucus)
L, > B
A, >|Direct contact| M[Cohabitation]
M, > C
N[Wild reservoirs: trout, herring, wild salmon], >|Shedding| B
O[Fomites, equipment], >|Mechanical vector| B
P[HPR0 endemic farms], >|Evolution via HPR deletion| Q[HPRΔ virulent variant]
Q, > A
FAQ
What is infectious salmon anaemia virus (ISAV)?
ISAV is an enveloped, negative-sense single-stranded RNA virus in the family Orthomyxoviridae, genus Isavirus, that causes a severe systemic disease in farmed Atlantic salmon [1].
What are the differences between HPR0 and HPRΔ variants of ISAV?
HPR0 variants possess a full-length highly polymorphic region in the hemagglutinin-esterase gene and are considered non-virulent, while HPRΔ variants have deletions in that region and are virulent [1, 31].
How is ISAV transmitted between fish?
ISAV is transmitted horizontally through water, direct contact, and possibly via fomites; vertical transmission via eggs is considered negligible under proper biosecurity [17, 19, 25].
Which fish species are susceptible to ISAV?
Atlantic salmon is the primary susceptible species, but experimental infections have shown susceptibility in rainbow trout, coho salmon, sea trout, and herring; lumpfish are refractory to infection [14, 11, 13, 16, 35].
What diagnostic methods are used for ISAV detection?
Real-time RT-PCR is the primary screening method, with confirmatory sequencing of segments 5 and 6 for genotyping; virus isolation in SHK-1 or ASK cells is used for characterization [21, 15, 20, 27].
Can ISAV be detected in water samples?
Yes, ISAV can be concentrated from seawater using charged membrane filters followed by RT-qPCR, enabling non-lethal surveillance of farm sites [26].
What cells are targeted by virulent ISAV?
Virulent ISAV targets endothelial cells lining blood vessels and the heart, causing systemic infection and anaemia; erythrocytes bind virus but do not support high-level replication [9, 12].
Does ISAV cause ocular pathologies in Atlantic salmon?
Yes, ISAV infection leads to corneal dysplasia, edema, and changes in the choroid rete mirabile, with accumulation of viral antigen in ocular tissues and dysregulation of ocular genes [10].
Is there a vaccine available for ISA?
Inactivated and recombinant vaccines exist but provide only partial protection; therefore, stamping out and biosecurity remain critical control measures [1].
How does HPR0 evolve into HPRΔ?
The transition involves intra-segmental recombination (template switching) in the HPR region of segment 6, resulting in deletions; additional mutations in segment 5 are required for full virulence [1, 31].
References
[1] Rimstad E, Markussen T. Infectious salmon anaemia virus, molecular biology and pathogenesis of the infection. Journal of Applied Microbiology. 2019. URL: https://www.semanticscholar.org/paper/5759df8c9e4eaae6be1dcb9ce55799e255e08fce
[2] Romero JF, Gardner I, Hammell L, et al. Descriptive epidemiology of variants of infectious salmon anaemia virus in four Atlantic salmon farms in Newfoundland and Labrador, Canada. Journal of Fish Diseases. 2022. URL: https://www.semanticscholar.org/paper/10c2fb798558141e34481a8229947eb7075d92e0
[3] Díaz A, García K, Navarrete A, et al. Virtual screening of gene expression regulatory sites in non-coding regions of the infectious salmon anemia virus. BMC Research Notes. 2014. URL: https://www.semanticscholar.org/paper/c25ec71a07bea2d0c9f19b0c9a4f91ba55cae907
[4] Ramly R, Olsen CM, Braaen S, et al. Infectious salmon anaemia virus nuclear export protein is encoded by a spliced gene product of genomic segment 7. Virus Research. 2013. URL: https://www.semanticscholar.org/paper/76057ea7ecd826b056cd55ee7c5678f5bceb574e
[5] Patel S, Dale OB, Spilsberg B, et al. A standardized bath challenge of Atlantic salmon reveals distinct infection dynamics and mortality across ten HPR-deleted infectious salmon anaemia virus isolates. bioRxiv. 2026. URL: https://www.semanticscholar.org/paper/fd411457c9a50478a822477b0c0c0685d4ea4884
[6] Christian RW. Morphology and morphogenesis of infectious salmon anaemia virus replicating in the endothelium of Atlantic salmon Salmo salar. Journal. Year unknown. URL: https://www.semanticscholar.org/paper/eb30f30c976ccb50dceadc558d3826e7ebf6e14e
[7] Tarnok ME, Gúzman F, Aguilar L. Effects of cholesterol on lipid vesicle fusion mediated by infectious salmon anaemia virus fusion peptides. Colloids and Surfaces B: Biointerfaces. 2022. URL: https://www.semanticscholar.org/paper/0bc7f1b54f1c42fe45294d27a9b0b047b5fd3a5c
[8] Cárdenas M, Michelson S, Galleguillos C, et al. Modulation of infectious Salmon Anaemia virus infection by clathrin-mediated endocytosis and macropinocytosis inhibitors. Research in Veterinary Science. 2024. URL: https://www.semanticscholar.org/paper/bf3ace5af0b4482fda268519f3b0d4922fbe793f
[9] Andresen AM, Taylor RS, Furniss JJ, et al. Virus-inclusive single-nucleus RNA sequencing reveals two distinct endothelial response patterns in infectious salmon anaemia. bioRxiv. 2025. URL: https://www.semanticscholar.org/paper/55c264560ac083f04c6c7862cdd1dd09ee8d6f80
[10] Mahon E, Kwabiah RR, Paradis H, et al. Ocular Manifestations of Infectious Salmon Anaemia Virus Infection in Atlantic Salmon (Salmo salar). Journal of Fish Diseases. 2026. URL: https://www.semanticscholar.org/paper/7b85f07c89118943912621eb3e94d15ea3e99b83
[11] MacWilliams C, Johnson G, Groman D, et al. Morphologic description of infectious salmon anaemia virus (ISAV)-induced lesions in rainbow trout Oncorhynchus mykiss compared to Atlantic salmon Salmo salar. Diseases of Aquatic Organisms. 2007. URL: https://www.semanticscholar.org/paper/e16e7674c9030d0b72034685c58b29133f1a8db2
[12] Fosse J, Aamelfot M, Sønstevold T, et al. Salmon Erythrocytes Sequester Active Virus Particles in Infectious Salmon Anaemia. Viruses. 2022. URL: https://www.semanticscholar.org/paper/7d07962dbbf3f546766b524238b09acbf3c39a54
[13] Nylund A, Devold M, Mullins J, et al. Herring (Clupea harengus): a host for infectious salmon anemia virus (ISAV). Journal. 2002. URL: https://www.semanticscholar.org/paper/164f091e5b81ef605a3946e369a14493d095328c
[14] Kibenge F, Gárate ON, Johnson G, et al. Isolation and identification of infectious salmon anaemia virus (ISAV) from Coho salmon in Chile. Diseases of Aquatic Organisms. 2001. URL: https://www.semanticscholar.org/paper/a4e8f846a2788c61fe9bc9e22f000e8ff800f077
[15] Devold M, Krossøy B, Aspehaug V, et al. Use of RT-PCR for diagnosis of infectious salmon anaemia virus (ISAV) in carrier sea trout Salmo trutta after experimental infection. Diseases of Aquatic Organisms. 2000. URL: https://www.semanticscholar.org/paper/e19dfa8dabcb6bbc84f4f1b631bffb96fb204df3
[16] Gagné N, Ditlecadet D, Hebert K, et al. Lack of Lumpfish (Cyclopterus lumpus) Susceptibility to the Infectious Salmon Anaemia Virus (ISAV) Using In Vivo Challenges. Journal of Fish Diseases. 2026. URL: https://www.semanticscholar.org/paper/2776dd23e711cc7c204b9d1778a1e90ec6ed3c14
[17] Nylund A, Plarre H, Karlsen M, et al. Transmission of infectious salmon anaemia virus (ISAV) in farmed populations of Atlantic salmon (Salmo salar). Archives of Virology. 2006. URL: https://www.semanticscholar.org/paper/7f19ab9842229bceae96f39b23f80effe6c120e2
[18] Weli S, Amundsen M, Dale OB, et al. Characterization of infectious salmon anaemia virus survival and infectivity in seawater. Aquaculture Reports. 2025. URL: https://www.semanticscholar.org/paper/d9d013a3792d51eb008a9759503e43fd8790a615
[19] Nielsen SS, Álvarez J, Boklund A, et al. Viral haemorrhagic septicaemia virus, infectious haematopoietic necrosis virus and HPR‑deleted infectious salmon anaemia virus – Risk of introduction in free areas through fertilised eggs and gametes. EFSA Journal. 2025. URL: https://www.semanticscholar.org/paper/54e183c46f627a81032b381cb5a9d96c04949af2
[20] Delphino MKVC, O'Brien N, Laurin E, et al. Bayesian analysis of diagnostic sensitivity and specificity for detecting infectious salmon anaemia virus (ISAV) using IFAT and real-time RT-PCR testing from laboratories in Atlantic Canada. Aquaculture. 2022. URL: https://www.semanticscholar.org/paper/f5bbb677081c22d1085e297
[21] Wergeland H, Jakobsen RA. A salmonid cell line (TO) for production of infectious salmon anaemia virus (ISAV). Diseases of Aquatic Organisms. 2001. URL: https://www.semanticscholar.org/paper/d02cfca730ba917d5692fe6ae22905c7d9ba8354
[22] Gervais O, Gratacap R, Papadopoulou A, et al. Understanding host response to infectious salmon anaemia virus in an Atlantic salmon cell line using single-cell RNA sequencing. bioRxiv. 2022. URL: https://www.semanticscholar.org/paper/940702ab743fa5b8d5221e97c40c82b46c32a0ca
[23] Wang J, Ballantyne M, Wang J, et al. WITHDRAWN: Triple gene disruption confers genetic resistance to infectious salmon anaemia virus in Atlantic salmon (Salmo salar). bioRxiv. 2025. URL: https://www.semanticscholar.org/paper/af488e7b31532284cf900ae2ce2203c8efbd8bda
[24] McBeath A, Aamelfot M, Christiansen D, et al. Immersion challenge with low and highly virulent infectious salmon anaemia virus reveals different pathogenesis in Atlantic salmon, Salmo salar L. Journal of Fish Diseases. 2015. URL: https://www.semanticscholar.org/paper/e2f5fdaff538e6c11c5d23b775bf8889595b1533