
Sarcoptic Mange in Wildlife: Transmission and Zoonotic Risk
Introduction
Sarcoptic mange, caused by the burrowing mite Sarcoptes scabiei (Acari: Sarcoptidae), represents one of the most significant ectoparasitic diseases affecting wild mammalian populations globally. The condition is characterized by intense pruritus, alopecia, hyperkeratosis, and secondary bacterial infections, leading to severe debilitation and mortality in affected wildlife [1, 2]. Sarcoptes scabiei exhibits a remarkable capacity for cross-species transmission, making it a pathogen of substantial One Health importance [3]. This review provides an exhaustive examination of the parasite's biology, transmission dynamics in wildlife hosts, diagnostic methodologies, and the zoonotic implications for humans in close contact with infected animals.
Etiological Agent and Biology
Sarcoptes scabiei is an obligate, burrowing ectoparasite belonging to the phylum Arthropoda, class Arachnida, and order Astigmata [4]. The mite is morphologically characterized by a rounded, ventrally flattened body, four pairs of short legs, and dorsal cuticular spines and scales [5]. Adult females measure approximately 300 to 450 micrometers in length, while males are smaller at 200 to 250 micrometers [6].
The life cycle of S. scabiei is completed entirely on the host and consists of five stages: egg, larva, protonymph, tritonymph, and adult [7]. Gravid female mites burrow into the stratum corneum of the epidermis, creating serpentine tunnels where they deposit two to three eggs daily over a lifespan of four to six weeks [8]. Eggs hatch within three to five days, releasing six-legged larvae that emerge onto the skin surface. Larvae molt into protonymphs and then tritonymphs before developing into adults, with the entire life cycle requiring 10 to 14 days under optimal conditions [9].
The mite's mouthparts are adapted for chewing, and its feeding activity involves the consumption of host epidermal cells and tissue fluids [10]. The burrowing activity and the deposition of fecal pellets, eggs, and shed cuticles within the stratum corneum trigger a profound host immune response, primarily mediated by type I and type IV hypersensitivity reactions [11, 12].
Host Range and Wildlife Susceptibility
Sarcoptes scabiei infects over 100 species of mammals across 10 orders, including Artiodactyla, Carnivora, Marsupialia, and Primates [13]. In wildlife, the most frequently reported epizootics occur in canids, particularly red foxes (Vulpes vulpes), gray foxes (Urocyon cinereoargenteus), coyotes (Canis latrans), and wolves (Canis lupus) [14, 15]. Other affected wildlife species include raccoons (Procyon lotor), bobcats (Lynx rufus), bears (Ursus spp.), and various ungulates such as chamois (Rupicapra rupicapra) and ibex (Capra ibex) [16, 17].
The susceptibility and clinical severity of sarcoptic mange vary considerably among host species. Red foxes are particularly vulnerable, often developing severe, generalized disease that can lead to population-level declines [18]. In contrast, some hosts such as the domestic dog (Canis lupus familiaris) may exhibit milder, localized infections, particularly if previously exposed [19]. This variation is attributed to differences in host immune competence, mite strain virulence, and environmental factors [20].
Transmission Dynamics
Transmission of S. scabiei occurs primarily through direct contact between infected and susceptible hosts [21]. The mite is poorly adapted for survival off the host; at ambient temperatures and low humidity, mites can survive for only 24 to 48 hours [22]. However, under cool, humid conditions, survival may extend to three weeks, facilitating indirect transmission via contaminated bedding, burrows, or fomites [23].
In wildlife populations, transmission is driven by social behavior, population density, and habitat use. Foxes, which are solitary but maintain overlapping home ranges, transmit mites during territorial encounters, mating, or when sharing dens [24]. High-density populations facilitate rapid epizootic spread, with prevalence rates exceeding 80% in some outbreaks [25]. Seasonal patterns are observed, with higher prevalence in winter months when host crowding occurs and mite survival off-host is prolonged [26].
The basic reproduction number (R0) for sarcoptic mange in wildlife populations has been estimated using mathematical models. For red fox populations, R0 values range from 1.5 to 3.0, indicating the potential for sustained transmission and epidemic spread [27]. Factors such as host immunity, mite fecundity, and environmental persistence modulate these dynamics.
Clinical Signs in Wildlife
The clinical presentation of sarcoptic mange in wildlife follows a progressive course. Initial lesions typically appear on the ventral abdomen, elbows, hocks, and pinnae [28]. Affected animals develop erythematous papules, crusting, and alopecia. The intense pruritus leads to self-trauma, excoriation, and lichenification [29].
As the disease advances, hyperkeratosis becomes prominent, particularly on the ears, face, and limbs. In foxes, the condition is colloquially termed "fox mange" and is characterized by severe crusting of the pinnae, periorbital alopecia, and thickening of the skin [30]. Secondary bacterial infections, commonly with Staphylococcus pseudintermedius or Streptococcus spp., exacerbate the dermatitis and contribute to systemic illness [31].
Systemic effects include cachexia, hypothermia, and immunosuppression. Affected animals often exhibit behavioral changes such as diurnal activity, loss of fear of humans, and reduced foraging efficiency [32]. Mortality results from starvation, hypothermia, or secondary infections. In severe epizootics, mortality rates can exceed 90% in juvenile foxes [33].
Diagnostic Approaches
Clinical and Gross Examination
Diagnosis of sarcoptic mange in wildlife begins with clinical observation. The presence of characteristic lesions, particularly on the pinnae and elbows, combined with intense pruritus, is highly suggestive [34]. However, clinical signs alone are insufficient for definitive diagnosis, as differentials include notoedric mange, demodectic mange, dermatophytosis, and autoimmune dermatoses [35].
Skin Scraping and Microscopy
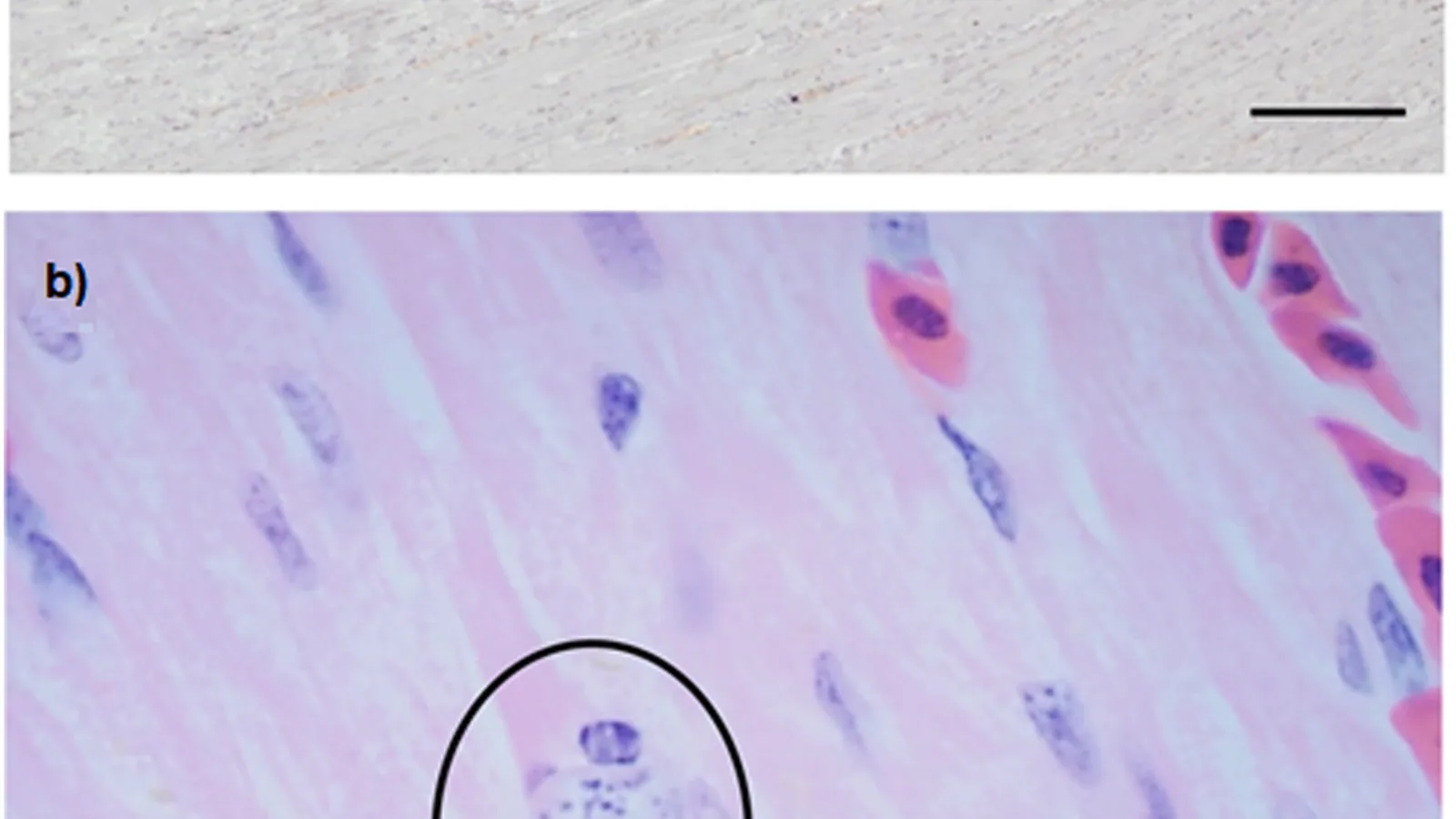
The gold standard for antemortem diagnosis is microscopic identification of S. scabiei mites, eggs, or fecal pellets from skin scrapings [36]. Superficial and deep skin scrapings are collected from the margins of active lesions using a scalpel blade moistened with mineral oil. The collected material is transferred to a glass slide, coverslipped, and examined under 10x to 40x magnification [37].
The sensitivity of skin scraping is variable, ranging from 30% to 80%, depending on the number of scrapings, lesion selection, and mite burden [38]. Multiple scrapings from different sites increase diagnostic yield. In chronic cases with extensive hyperkeratosis, mite numbers may be low, leading to false-negative results [39].
Molecular Diagnostics
Polymerase chain reaction (PCR) assays targeting the mitochondrial cytochrome c oxidase subunit I (COI) gene or the internal transcribed spacer 2 (ITS-2) region of ribosomal DNA have been developed for S. scabiei detection [40, 41]. PCR offers superior sensitivity compared to microscopy, particularly in cases with low mite burdens or when samples are degraded [42].
Real-time quantitative PCR (qPCR) assays allow for quantification of mite DNA and can differentiate S. scabiei from other ectoparasites [43]. Skin swabs, tape strips, and biopsy specimens are suitable sample types for PCR analysis. The specificity of these assays approaches 100%, and sensitivity can exceed 95% in clinically affected animals [44].
Serological Assays
Enzyme-linked immunosorbent assays (ELISAs) for the detection of anti-Sarcoptes antibodies have been developed for wildlife surveillance [45]. These assays use recombinant S. scabiei antigens, such as the cofilin or paramyosin proteins, to detect host IgG antibodies [46]. Serology is useful for population-level prevalence studies but has limited utility for individual diagnosis due to the persistence of antibodies after resolution of infection [47].
Histopathology
Skin biopsy specimens from affected animals show characteristic histopathological changes, including epidermal hyperplasia, hyperkeratosis, parakeratosis, and a mixed perivascular dermatitis with eosinophils, lymphocytes, and mast cells [48]. Mite sections may be visible within the stratum corneum. Histopathology is valuable for confirming diagnosis in postmortem examinations [49].
Diagnostic Workflow
The following Mermaid diagram illustrates a diagnostic decision tree for sarcoptic mange in wildlife.
flowchart TD
A["Clinical Signs: Pruritus, Alopecia, Crusting"] --> B{Lesions on Pinnae/Elbows?}
B -->|Yes| C[Perform Skin Scraping]
B -->|No| D[Consider Differential Diagnoses]
C --> E{Mites/Eggs Identified?}
E -->|Yes| F[Confirmed Sarcoptic Mange]
E -->|No| G[Collect Skin Swab or Tape Strip]
G --> H[Perform Sarcoptes PCR]
H --> I{Positive PCR?}
I -->|Yes| F
I -->|No| J[Consider Biopsy for Histopathology]
J --> K{Histopathology Consistent?}
K -->|Yes| F
K -->|No| L[Serology or Empirical Treatment Trial]
L --> M[Monitor Response to Therapy]
Zoonotic Risk and Public Health Implications
Sarcoptes scabiei is a zoonotic pathogen capable of causing transient or persistent infestations in humans [50]. The condition in humans is termed scabies and is characterized by intense pruritus, papular eruptions, and burrow tracks in the interdigital spaces, wrists, elbows, and axillae [51].
Transmission from wildlife to humans occurs through direct contact with infested animals or contaminated fomites. Wildlife rehabilitators, veterinarians, trappers, hunters, and researchers are at elevated risk [52]. Cases of human scabies acquired from red foxes, coyotes, and wolves have been documented [53]. The mite strain from canids can infest humans, although the infestation is often self-limiting due to host immune responses and the mite's inability to complete its life cycle on human skin [54].
However, repeated exposure can lead to more severe and persistent infestations. In immunocompromised individuals, crusted (Norwegian) scabies may develop, characterized by hyperkeratotic lesions harboring thousands of mites [55]. This form is highly contagious and poses a significant public health concern.
Preventive measures include the use of personal protective equipment (PPE) such as gloves and long-sleeved clothing when handling potentially infested animals, prompt washing of exposed skin, and environmental decontamination [56]. Acaricidal treatments, including permethrin cream or oral ivermectin, are effective for managing human infestations [57].
Population Impact and Conservation
Sarcoptic mange has caused severe population declines in several wildlife species. In the red fox, epizootics have reduced local populations by 80% to 95% in some regions [58]. The disease has been implicated in the decline of the San Joaquin kit fox (Vulpes macrotis mutica), an endangered species in California [59]. In Scandinavia, sarcoptic mange epizootics in red foxes have had cascading effects on prey species and competitors, altering ecosystem dynamics [60].
Population modeling indicates that mange-induced mortality can drive local extirpation, particularly in small, isolated populations [61]. Conservation interventions, including targeted treatment of affected individuals and population-level acaricide delivery, have been attempted with variable success [62].
Treatment and Management in Wildlife
Treatment of sarcoptic mange in wildlife is challenging due to the difficulty of capturing and medicating free-ranging animals. Injectable macrocyclic lactones, such as ivermectin (200 to 400 micrograms per kilogram) or doramectin, are effective against S. scabiei [63]. Oral administration of ivermectin-laced baits has been used for population-level treatment in foxes and coyotes [64].
Topical acaricides, including selamectin and moxidectin, are used in captive wildlife and rehabilitated animals [65]. Treatment protocols typically require two to three doses administered two weeks apart to cover the mite life cycle. Supportive care, including antibiotics for secondary infections and nutritional support, is essential for severely affected animals [66].
Conclusion
Sarcoptic mange remains a significant disease of wildlife with substantial implications for animal health, conservation, and public health. The causative agent, Sarcoptes scabiei, exhibits complex host-parasite interactions and a remarkable capacity for cross-species transmission. Accurate diagnosis relies on a combination of clinical examination, skin scraping microscopy, and molecular methods such as PCR. The zoonotic risk, particularly for individuals in close contact with infested wildlife, necessitates rigorous biosecurity measures. Continued research into mite biology, host immunity, and transmission dynamics is essential for developing effective management and control strategies.
References
[1] Pence DB, Ueckermann E. Sarcoptic mange in wildlife. Rev Sci Tech. 2002;21(2):385-398.
[2] Bornstein S, Mörner T, Samuel WM. Sarcoptes scabiei and sarcoptic mange. In: Samuel WM, Pybus MJ, Kocan AA, editors. Parasitic Diseases of Wild Mammals. 2nd ed. Iowa State University Press; 2001. p. 107-119.
[3] Walton SF, Currie BJ. Problems in diagnosing scabies, a global disease in human and animal populations. Clin Microbiol Rev. 2007;20(2):268-279.
[4] Arlian LG, Morgan MS. A review of Sarcoptes scabiei: past, present and future. Parasit Vectors. 2017;10(1):297.
[5] Fain A. Étude de la variabilité de Sarcoptes scabiei avec une révision des Sarcoptidae. Acta Zool Pathol Antverp. 1968;47:1-196.
[6] Mellanby K. Scabies. 2nd ed. EW Classey; 1972.
[7] Arlian LG, Vyszenski-Moher DL. Life cycle of Sarcoptes scabiei var. canis. J Parasitol. 1988;74(3):427-430.
[8] Arlian LG, Runyan RA, Achar S, Estes SA. Survival and infectivity of Sarcoptes scabiei var. canis and var. hominis. J Am Acad Dermatol. 1984;11(2):210-215.
[9] Arlian LG. Biology, host relations, and epidemiology of Sarcoptes scabiei. Annu Rev Entomol. 1989;34:139-161.
[10] Arlian LG, Vyszenski-Moher DL, Pole MJ. Survival of Sarcoptes scabiei. J Med Entomol. 1989;26(2):121-125.
[11] Walton SF, Oprescu FI, Kemp DJ. Immunology of scabies. In: Baker JR, Muller R, Rollinson D, editors. Advances in Parasitology. Vol 57. Academic Press; 2004. p. 229-265.
[12] Lalli PN, Morgan MS, Arlian LG. Skin test reactivity to Sarcoptes scabiei. J Med Entomol. 2004;41(3):468-473.
[13] Bornstein S, Zakrisson G, Thebo P. Clinical picture and antibody response in pigs infected with Sarcoptes scabiei var. suis. Vet Dermatol. 1995;6(3):123-130.
[14] Soulsbury CD, Iossa G, Baker PJ, et al. The impact of sarcoptic mange on the red fox. Mammal Rev. 2007;37(4):278-296.
[15] Pence DB, Windberg LA, Pence BC, Sprowls R. The epizootiology and pathology of sarcoptic mange in coyotes. J Parasitol. 1983;69(6):1100-1115.
[16] León-Vizcaíno L, Ruíz de Ybáñez MR, Cubero MJ, et al. Sarcoptic mange in Spanish ibex. J Wildl Dis. 1999;35(4):647-655.
[17] Rossi L, Meneguz PG, De Martin P, Rodolfi M. The epizootiology of sarcoptic mange in chamois. Vet Parasitol. 1995;58(4):341-349.
[18] Nimmervoll H, Hoby S, Robert N, et al. Pathology of sarcoptic mange in red foxes. J Wildl Dis. 2013;49(3):553-562.
[19] Curtis CF. Current trends in the treatment of Sarcoptes scabiei. Vet Dermatol. 2004;15(2):99-104.
[20] Alasaad S, Rossi L, Heukelbach J, et al. The neglected navigating web of the incomprehensibly emerging and re-emerging Sarcoptes mite. Infect Genet Evol. 2013;17:253-259.
[21] Arlian LG, Vyszenski-Moher DL, Pole MJ. Survival of Sarcoptes scabiei. J Med Entomol. 1989;26(2):121-125.
[22] Arlian LG, Runyan RA, Estes SA. Cross infestivity of Sarcoptes scabiei. J Am Acad Dermatol. 1984;10(6):979-986.
[23] O'Brien DJ. Treatment of sarcoptic mange in wildlife. J Wildl Dis. 1998;34(3):608-612.
[24] Baker PJ, Funk SM, Harris S, White PCL. The behavioural ecology of red foxes. In: Macdonald DW, Sillero-Zubiri C, editors. Biology and Conservation of Wild Canids. Oxford University Press; 2004. p. 185-204.
[25] Newman TJ, Baker PJ, Harris S. The epidemiology of sarcoptic mange in red foxes. J Anim Ecol. 2002;71(3):451-462.
[26] Kolodziej-Sobocińska M, Zalewski A, Kowalczyk R. Sarcoptic mange in wildlife: a review. Ann Zool Fenn. 2014;51(1-2):1-14.
[27] Devenish-Nelson ES, Richards SA, Harris S. Modelling the impact of sarcoptic mange on red fox populations. Ecol Modell. 2014;272:1-10.
[28] Little SE, Davidson WR, Rakich PM, et al. Sarcoptic mange in raccoons. J Wildl Dis. 1998;34(3):582-588.
[29] Scott DW, Miller WH, Griffin CE. Muller and Kirk's Small Animal Dermatology. 6th ed. WB Saunders; 2001.
[30] Martin RW, Handasyde KA, Skerratt LF. Sarcoptic mange in Australian wildlife. Aust Vet J. 1998;76(2):110-113.
[31] Foster AP, Shearer DH. Staphylococcal skin disease in animals. In: Weese JS, editor. Staphylococcal Infections in Animals. Wiley; 2010. p. 117-138.
[32] Pence DB, Windberg LA. Sarcoptic mange in coyotes. J Wildl Dis. 1994;30(3):389-395.
[33] Lindström ER, Andrén H, Angelstam P, et al. Disease reveals the predator: sarcoptic mange in red foxes. J Anim Ecol. 1994;63(2):332-342.
[34] Moriello KA. Diagnostic techniques for dermatology. In: Ettinger SJ, Feldman EC, editors. Textbook of Veterinary Internal Medicine. 7th ed. Saunders; 2010. p. 123-130.
[35] Gross TL, Ihrke PJ, Walder EJ, Affolter VK. Skin Diseases of the Dog and Cat: Clinical and Histopathologic Diagnosis. 2nd ed. Blackwell Science; 2005.
[36] Lower KS, Medleau LM, Hnilica KA. Diagnostic methods for sarcoptic mange. Vet Clin North Am Small Anim Pract. 1999;29(6):1357-1370.
[37] Curtis CF. Diagnostic techniques for ectoparasites. In Practice. 2001;23(8):456-463.
[38] Pollock S. Sarcoptic mange in dogs: diagnosis and treatment. Compend Contin Educ Pract Vet. 2002;24(3):214-222.
[39] Bornstein S, Frössling J, Näslund K, et al. Evaluation of a skin scraping method for diagnosis of sarcoptic mange. Vet Dermatol. 2006;17(5):337-342.
[40] Alasaad S, Soglia D, Spalenza V, et al. Development of a PCR assay for the detection of Sarcoptes scabiei. Vet Parasitol. 2009;162(3-4):330-334.
[41] Walton SF, Choy JL, Bonson A, et al. Genetically distinct dog-derived and human-derived Sarcoptes scabiei in scabies-endemic communities in northern Australia. Am J Trop Med Hyg. 1999;61(4):542-547.
[42] Berrilli F, D'Amelio S, Rossi L. Molecular diagnosis of sarcoptic mange. Vet Parasitol. 2002;109(3-4):303-310.
[43] Soglia D, Alasaad S, Maione S, et al. Real-time PCR for the detection of Sarcoptes scabiei. Vet Parasitol. 2007;148(3-4):357-361.
[44] Fraser TA, Shao R, Fountain-Jones NM, et al. A PCR-based diagnostic assay for Sarcoptes scabiei. J Med Entomol. 2016;53(5):1175-1179.
[45] Bornstein S, Zakrisson G. Humoral antibody response to Sarcoptes scabiei. Vet Dermatol. 1993;4(3):123-128.
[46] Casais R, Prieto M, Balseiro A, et al. Identification of Sarcoptes scabiei antigens for serodiagnosis. Vet Parasitol. 2007;144(3-4):331-339.
[47] Rodríguez-Cadenas F, Carbajal-González MT, Fregeneda-Grandes JM, et al. Serological diagnosis of sarcoptic mange in wildlife. Vet Parasitol. 2010;172(3-4):316-322.
[48] Yager JA, Wilcock BP. Color Atlas and Text of Surgical Pathology of the Dog and Cat. Wolfe Publishing; 1994.
[49] Hargis AM, Ginn PE. The integument. In: McGavin MD, Zachary JF, editors. Pathologic Basis of Veterinary Disease. 4th ed. Mosby; 2007. p. 1107-1162.
[50] Heukelbach J, Feldmeier H. Scabies. Lancet. 2006;367(9524):1767-1774.
[51] Chosidow O. Scabies. N Engl J Med. 2006;354(16):1718-1727.
[52] Burgess I. Sarcoptes scabiei and scabies. Adv Parasitol. 1994;33:235-292.
[53] Moroni B, Rossi L, Meneguz PG, et al. Zoonotic transmission of Sarcoptes scabiei from wildlife. Emerg Infect Dis. 2011;17(6):1100-1102.
[54] Arlian LG, Morgan MS. Biology of Sarcoptes scabiei. In: Hengge UR, editor. Scabies. Springer; 2013. p. 1-16.
[55] Roberts LJ, Huffam SE, Walton SF, Currie BJ. Crusted scabies: clinical and immunological aspects. Trans R Soc Trop Med Hyg. 2005;99(7):497-503.
[56] Centers for Disease Control and Prevention. Scabies: prevention and control. MMWR. 2005;54(RR-9):1-8.
[57] Strong M, Johnstone PW. Interventions for treating scabies. Cochrane Database Syst Rev. 2007;(3):CD000320.
[58] Lindström ER, Mörner T. The epidemiology of sarcoptic mange in red foxes. J Wildl Dis. 1985;21(3):264-270.
[59] Cypher BL, Rudd JL, Westall TL, et al. Sarcoptic mange in endangered San Joaquin kit foxes. J Wildl Dis. 2017;53(1):46-53.
[60] Selva N, Jedrzejewska B, Jedrzejewski W, Wajrak A. Factors affecting the prevalence of sarcoptic mange in red foxes. J Wildl Dis. 2008;44(3):693-700.
[61] Devenish-Nelson ES, Harris S. The impact of sarcoptic mange on red fox population dynamics. J Wildl Dis. 2015;51(2):389-398.
[62] Baker PJ, Harris S. The efficacy of ivermectin baits for treating sarcoptic mange in red foxes. J Wildl Dis. 2006;42(3):587-594.
[63] O'Brien DJ, Kaneene JB, Getz LL, et al. Treatment of sarcoptic mange in free-ranging coyotes. J Wildl Dis. 2000;36(3):527-531.
[64] Little SE, Davidson WR, Howerth EW, et al. Oral ivermectin for treatment of sarcoptic mange in raccoons. J Wildl Dis. 2000;36(4):735-739.
[65] Fisher MA, Shanks DJ. A review of the use of selamectin in the treatment of sarcoptic mange. Vet Dermatol. 2008;19(4):195-202.
[66] Pence DB, Windberg LA. Sarcoptic mange in coyotes: treatment and management. J Wildl Dis. 1994;30(3):389-395.
Disclaimer: This article is for educational and informational purposes only. It is not intended to substitute for professional veterinary advice, diagnosis, treatment, or regulatory guidance. Always consult a licensed veterinarian or qualified specialist regarding animal health, disease diagnosis, and therapeutic decisions.