
Thelazia (Eyeworm): Taxonomy, Epidemiology, Clinical Management, and Vector Biology
Thelazia species (Spirurida: Thelaziidae), commonly known as eyeworms, are parasitic nematodes that inhabit the conjunctival sac, lacrimal ducts, and orbital cavities of a wide range of vertebrate hosts, including domestic carnivores, wild mammals, livestock, birds, and humans [1, 2, 3]. These nematodes are transmitted by drosophilid flies of the genus Phortica (subfamily Steganinae), which act as obligate intermediate hosts and vectors [2, 4, 5]. Ocular thelaziosis has emerged as a significant veterinary concern across Europe, Asia, and increasingly in the Americas, driven by sylvatic reservoir hosts and expanding vector distributions [6, 7, 8, 9, 24].
Taxonomy and Species Diversity
The genus Thelazia is divided into two subgenera: Thelazia sensu stricto (parasitizing mammals) and Thelaziella (parasitizing birds) [10]. The most medically and veterinary relevant mammalian species are summarized in Table 1.
Table 1. Major Thelazia species of veterinary importance.
| Species | Primary Hosts | Geographic Distribution | Vector (known or suspected) |
|---|---|---|---|
| Thelazia callipaeda | Canids, felids, mustelids, ursids, lagomorphs, humans | Europe, Asia, North America (emerging) | Phortica variegata, Phortica okadai |
| Thelazia rhodesi | Cattle, bison, other ruminants | Africa, Asia, Europe | Muscid flies (suspected) |
| Thelazia gulosa | Cattle, bison | North America, Europe, Asia | Muscid flies (suspected) |
| Thelazia skrjabini | Cattle, bison, sheep | Europe, Asia | Muscid flies (suspected) |
| Thelazia lacrymalis | Horses, camels | Asia, Europe | Muscid flies (suspected) |
| Thelazia leesei | Dromedary camels, other camelids | Middle East, Asia | Unknown |
| Thelazia aquilina (subgenus Thelaziella) | Birds of prey (e.g., Harpia harpyja) | South America, Africa | Unknown |
Thelazia callipaeda (the "oriental eyeworm") is the most intensively studied species due to its zoonotic potential and expanding range [2, 4, 8, 9]. Molecular characterization of the cytochrome c oxidase subunit I (cox1) gene has identified multiple haplotypes; haplotype 1 (h1) is the predominant lineage circulating in Europe and North America [4, 7, 11, 9]. Two unique lineages have been reported in Japan, suggesting endemic diversification [9].
Morphology
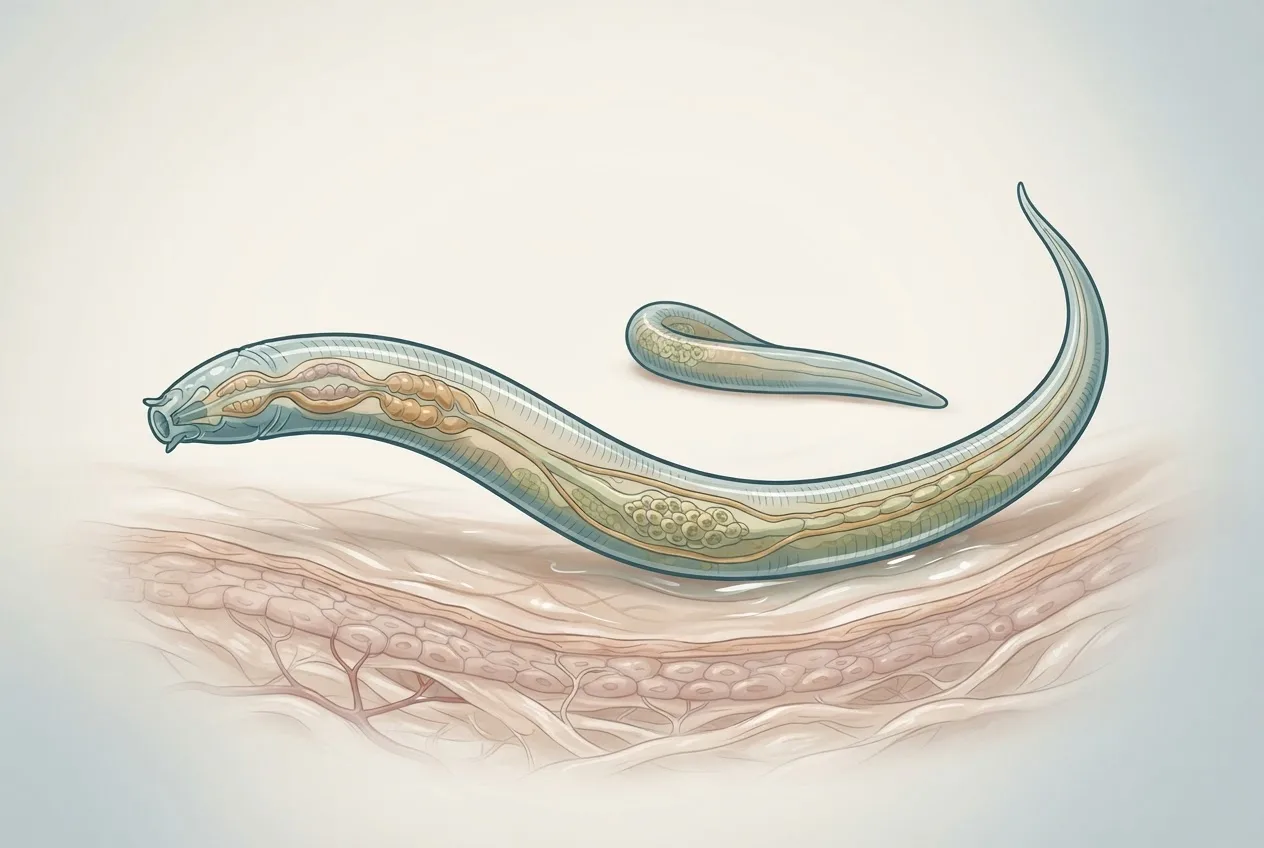
Adult Thelazia callipaeda are small, whitish nematodes. Males measure 7.7–11.9 mm in length and 300–400 µm in width; females are larger (12–19 mm by 350–500 µm) [3, 24]. The anterior end bears a characteristic scalariform (ladder-like) buccal capsule, a short muscular esophagus, and prominent cervical papillae [3, 28]. Females have a vulva located anterior to the esophago-intestinal junction, which is a key diagnostic feature [3]. The cuticle exhibits fine transverse striations. Scanning electron microscopy of the related Phortica okadai vector reveals five types of head sensilla (chaetica, basiconic, coeloconic, trichoid, and campaniform) with sexual dimorphism in the length of certain subtypes [5].
Life Cycle and Vector Biology
The life cycle of Thelazia callipaeda is indirect and obligately involves dipteran vectors of the genus Phortica (family Drosophilidae) [2, 5, 21]. The complete developmental cycle is depicted in the Mermaid diagram below.
graph TD
A[Adult worms in mammalian eye], > B{Gravid female releases L1 larvae into lacrimal secretions}
B, > C[Phortica vector feeds on lachrymal fluid]
C, > D[L1 ingested by fly]
D, > E[L1 develop to L3 infective larvae in fly hemocoel]
E, > F[Fly feeds on another mammal]
F, > G[L3 deposited on conjunctiva]
G, > H[L3 develop to adults in conjunctival sac]
H, > A
E, > I[L3 overwinter in vector?], > F
F, > J[Transmission season: spring-autumn in temperate regions]
Figure 1. Life cycle of Thelazia callipaeda.
Adult female Thelazia callipaeda deposit first-stage larvae (L1) into the lachrymal fluid of the definitive host [4]. These larvae are ingested by a lachryphagous Phortica fly during tear feeding [2]. Within the fly, L1 penetrate the gut wall and migrate to the hemocoel, where they molt twice to the infective third stage (L3) [21]. The developmental period within the vector is temperature dependent, typically requiring 14–21 days under field conditions [2, 5]. Infective L3 migrate to the fly's proboscis and are deposited onto the conjunctiva of a new host during subsequent tear-feeding events [2].
The principal European vector is Phortica variegata, which exhibits marked sexual dimorphism in feeding behavior: males are strongly lachryphagous and are the primary transmitters, whereas females feed predominantly on fruit and plant exudates [2, 21]. In Asia, Phortica okadai serves as the dominant vector, and its chromosome-level genome assembly has been published [32]. Recently, Phortica oldenbergi was reported in Italy for the first time, though it was not found infected with T. callipaeda [21]. Wolbachia pipientis endosymbionts have been detected in P. oldenbergi and P. okadai but not in P. variegata [21].
Olfactory ecology of P. variegata reveals higher antennal responses to phenol, 3-octanone, and sulcatone in males, suggesting a role in host seeking. The antennae respond more strongly to microbial and yeast-related volatiles (e.g., ethyl propanoate, butyl propanoate, nonanal) than to plant terpenoids, indicating that lure development should target these compounds [2].
Host Range and Epidemiology
Thelazia callipaeda infects a remarkably broad range of mammalian hosts. In Europe, wild carnivores serve as important reservoirs. Prevalence in red foxes (Vulpes vulpes) ranges from 3.9% (Croatia) to 12.2% (Hungary) and up to nearly 50% in some Portuguese studies [4, 24, 35]. Golden jackals (Canis aureus) show 4.3–9.7% prevalence in Croatia and Hungary, with a high proportion of gravid females (73.7% in jackals) indicating a key reservoir role [4, 35]. Iberian wolves (Canis lupus signatus) in northwestern Spain have an overall prevalence of 17%, with higher infection in young animals (1–3 years) and in regions with humid river basins (Miño and Sil) that favor vector development [7]. In contrast, stone martens (Martes foina) and European badgers (Meles meles) have not been found infected in Croatia or Hungary, although a Japanese marten (Martes melampus melampus) was infected [1, 4, 35].
In Japan, additional wild hosts include the Japanese red fox, masked palm civet (Paguma larvata), Japanese badger (Meles anakuma), Japanese black bear (Ursus thibetanus japonicus), raccoon (Procyon lotor), and raccoon dog (Nyctereutes viverrinus) [9]. The leopard cat (Prionailurus bengalensis) and raccoon dog (Nyctereutes procyonoides) were reported infected in Korea [3]. In North America, T. callipaeda has emerged in domestic dogs in the northeastern United States and was isolated from an American black bear (Ursus americanus) in Pennsylvania, confirming a sylvatic cycle [8, 12]. Autochthonous feline cases were also documented in the U.S. [8].
Cattle and bison are infected by T. rhodesi, T. gulosa, and T. skrjabini. In Ethiopian cattle, prevalence of T. rhodesi and T. gulosa reached 23.9%, with highest rates in autumn (39.7%) and in adult local breed animals under extensive management [6]. European bison (Bison bonasus) in Poland show a 61.9% prevalence (mainly T. skrjabini and T. gulosa), with a significant decrease in the Lowland genetic line over the study period [13]. Thelazia skrjabini was significantly associated with severity of ocular lesions [13]. Romanian bison (both wild European and farmed American) also harbored T. rhodesi and T. skrjabini [14]. Camels in Iran are infected with T. leesei (prevalence 29.5%), with higher infection in animals over 4 years old and significant association with lacrimation [28].
Seasonal dynamics vary by host. In red foxes, infection peaks in autumn and winter, coinciding with peak reproductive activity of the worm [4]. Golden jackals show more stable year-round infections, likely supporting continuous transmission [4]. Environmental drivers of vector abundance include temperature, wind speed, barometric pressure, and the presence of hygrophilous beech forests, which maintain high humidity [21, 35].
Clinical Signs and Pathogenesis
Ocular thelaziosis in dogs and cats commonly presents with epiphora, conjunctivitis (including follicular conjunctivitis), blepharospasm, uveitis, corneal ulceration, and keratitis [11, 25, 30]. In severe cases, corneal opacity and blindness may ensue [6]. However, asymptomatic infections are well documented; worms may be discovered incidentally during cataract surgery or routine ophthalmic examination [22, 23]. In cattle, lacrimation, conjunctivitis, and corneal ulcers are significantly associated with infection, and the presence of Thelazia spp. is positively associated with ocular lesion occurrence (p < 0.001) [6]. In European bison, T. skrjabini infection correlates with more severe ocular lesions, but the role of Thelazia in infectious keratoconjunctivitis (IKC) remains unclear; histopathology did not confirm a direct causal link, although prevalence was high (66.4%) [27].
Immunopathogenesis involves the secretion of macrophage migration inhibitory factor (T.cp-MIF) by T. callipaeda, which induces a dynamic M1-to-M2 macrophage polarization in THP-1-derived macrophages via the TLR4–NF-κB and PI3K/Akt signaling pathways [26, 33]. The C/EBPβ-SESN2 axis promotes M2b polarization, suppressing inflammation and potentially facilitating chronic infection [26].
Diagnosis
Definitive diagnosis relies on direct visualization and retrieval of adult worms from the conjunctival sac, third eyelid, or lacrimal ducts during ophthalmic examination. Worms are visible to the naked eye, but thorough inspection with lid eversion (double eversion) is essential to avoid missing small or larval stages [23, 30]. Morphological identification uses light microscopy to assess buccal capsule structure, esophageal length, and vulvar position [3, 28].
Molecular confirmation is achieved by PCR amplification and sequencing of the cox1 gene, which allows species identification and haplotype determination [4, 7, 11, 9, 24]. Conventional PCR using generic nematode primers targeting cox1 is the standard; real-time PCR assays have been developed but are not yet widely commercialized. Differentiation of the four bovine species (T. rhodesi, T. gulosa, T. skrjabini, T. lacrymalis) requires morphometric analysis and cox1 sequencing [14, 13, 27].
Treatment
Mechanical removal of adult worms from the eye remains the cornerstone of therapy [23, 30]. In companion animals, topical or systemic anthelmintics are used to kill residual worms. A topical combination of esafoxolaner, eprinomectin, and praziquantel (a macrocyclic lactone-based product applied to the skin) demonstrated 100% efficacy against T. callipaeda in naturally infected cats, with resolution of ocular signs within 14 days [25]. Systemic macrocyclic lactones (ivermectin, milbemycin oxime, moxidectin) are effective but are contraindicated in dogs with the MDR1 mutation (e.g., Collies); mechanical removal and local anti-inflammatory therapy provide an alternative [30]. In cattle, levamisole (15 mg/kg orally or by injection) and ivermectin (0.2–0.5 mg/kg) have been used [15, 31]. In vitro nematicidal effects of levamisole against T. callipaeda have been confirmed, and an experimental animal model for efficacy testing has been developed [15].
Control and Prevention
Vector control targeting Phortica spp. is challenging because adult flies are not attracted to standard traps used for muscid flies. Olfactory attractants identified from antennal studies (phenol, 3-octanone, nonanal, decanal) are under investigation for lure development [2]. In endemic areas, preventive use of monthly macrocyclic lactones in dogs and cats (especially those that travel) may reduce infection risk. Surveillance of wild carnivore populations is critical for early detection of emerging foci [4, 7, 24, 35]. The introduction of T. callipaeda to the Canary Islands via travel of infected dogs highlights the need for travel history assessment and border surveillance [11]. In North America, the recent autochthonous cases in dogs, cats, and a bear warrant heightened vigilance [8, 12].
Genomics
The nuclear genome of T. callipaeda has been assembled to a total length of 117.59 Mb, with 75.82% scaffolded into four chromosomal pseudomolecules including the X sex chromosome; the mitochondrial genome (13.66 kb) is also available [16]. The genome of its vector P. okadai (263.82 Mb) has been assembled at chromosome level (6 pseudo-chromosomes) with 15,052 protein-coding genes [32]. These resources facilitate studies on anthelmintic resistance, host-parasite coevolution, and vector competence.
Frequently Asked Questions
What is thelaziasis?
Thelaziasis is an ocular parasitic disease caused by nematodes of the genus Thelazia, characterized by infestation of the conjunctival sac and lacrimal ducts, leading to conjunctivitis, keratitis, and epiphora [1, 4, 6].
Which animals are most commonly affected by Thelazia callipaeda?
Dogs and cats are the most commonly reported domestic hosts, but wild carnivores such as red foxes, golden jackals, and wolves serve as major sylvatic reservoirs [4, 7, 24, 35].
How is Thelazia callipaeda transmitted?
Transmission occurs through the bite of lachryphagous drosophilid flies of the genus Phortica (primarily P. variegata in Europe and P. okadai in Asia) that deposit infective third-stage larvae into the eye during tear feeding [2, 5, 21].
What are the clinical signs of thelaziosis in dogs and cats?
Common signs include epiphora, conjunctivitis (including follicular), blepharospasm, uveitis, corneal ulcers, and in chronic cases, corneal opacity and blindness [11, 25, 30].
How is thelaziasis diagnosed?
Diagnosis is made by direct visualization of worms in the eye, followed by morphological identification (buccal capsule shape, vulvar position) and molecular confirmation via cox1 sequencing [3, 4, 24].
What treatments are available for Thelazia infection in dogs and cats?
Topical application of a combination of esafoxolaner, eprinomectin, and praziquantel (systemic macrocyclic lactone) showed 100% efficacy in cats [25]. Mechanical removal of worms and local anti-inflammatory therapy are also essential [30].
Can humans get infected with Thelazia callipaeda?
Yes, Thelazia callipaeda is zoonotic; human cases have been reported in Asia and Europe, often associated with rural living, contact with dogs, and presence of the vector [17, 18, 29].
How can ocular thelaziosis be prevented in pets?
Prevention in endemic areas involves regular use of macrocyclic lactone-based anthelmintics (e.g., milbemycin oxime, moxidectin) and minimizing exposure to Phortica flies by avoiding outdoor activities during peak vector activity (dusk/dawn) [2, 25].
What is the role of wildlife in thelaziosis epidemiology?
Wild carnivores, particularly foxes and jackals, act as reservoir hosts, maintaining transmission cycles and facilitating geographic expansion of the parasite [4, 7, 35]. In Japan, bears and martens also contribute [1, 9].
Is there a vaccine against Thelazia?
No vaccine is currently available. Control relies on chemotherapy, vector management, and surveillance [2, 25, 35].
References
[1] Tokiwa T, Doi K, Kitajima A. Wild Japanese Marten, Martes melampus melampus (Carnivora: Mustelidae), Is a New Host for Zoonotic Thelazia callipaeda Eyeworm. Ecology and Evolution. 2025. https://www.semanticscholar.org/paper/ecec9e2096ff217bae79ce8ff82944fc3a472a07
[2] Erdei AL, Szelényi M, Deutsch F, et al. Olfactory responses of the variegated fruit fly, Phortica variegata, an emerging vector of the zoonotic eyeworm Thelazia callipaeda, to ecologically relevant volatiles. Parasites & Vectors. 2025. https://www.semanticscholar.org/paper/1dd165599bd25f15acc98b5d6e7b4cc31f08857a
[3] Jang J, Park YS, Yun YM, et al. Thelazia callipaeda (Nematoda: Spirurida) infections in 2 wildlife species, Prionailurus bengalensis and Nyctereutes procyonoides, in Korea. Parasites, hosts and diseases. 2025. https://www.semanticscholar.org/paper/c235daf25820d37f27977bae46dc1ece0caba1a5
[4] Gagović E, Naletilić Š, Jurković Žilić D, et al. Eyes in the wild: Thelazia callipaeda in wild carnivores of Croatia. Parasites & Vectors. 2025. https://www.semanticscholar.org/paper/18f234ef23d528017c4081bf1cb4380ed1b3d925
[5] Sun D, Luo Y, Wang Y, et al. Ultrastructural Characterization of Developmental Stages and Head Sensilla in Portici okadai, Vector of Thelazia callipaeda. Insects. 2025. https://www.semanticscholar.org/paper/ffcae18ac68afed41ea1584d0e4fb982e5fdd309
[6] Alemneh T, Dagnachew S. Thelazia species (eyeworms) infection in cattle: Prevalence, species diversity, seasonal dynamics and its ocular effects in South Gondar zone, Northwest Ethiopia. Veterinary Parasitology: Regional Studies and Reports. 2025. https://www.semanticscholar.org/paper/437b9ad1c8eb52ff1d287831f3eb77598e8da8d7
[7] Estévez-Sánchez E, López-Beceiro A, González-Serrano C, et al. Thelazia callipaeda infection in Northwestern Spain: what role does the Iberian wolf play? International Journal for Parasitology: Parasites and Wildlife. 2025. https://www.semanticscholar.org/paper/b1ce955ff820724686380d5ad74c99afd9354190
[8] Manoj RRS, White H, Young R, et al. Emergence of Thelaziosis Caused by Thelazia callipaeda in Dogs and Cats, United States. Emerging Infectious Diseases. 2024. https://www.semanticscholar.org/paper/266f090111452245555ec94b29b291e685509e25
[9] Kitajima A, Tokiwa T, Doi K, et al. New host record of Thelazia callipaeda (Nematoda: Spirurida) with a notably wide host range and shared zoonotic lineage in Japan. Parasitology International. 2024. https://www.semanticscholar.org/paper/1d41de8ca4b018a4048af6591eb75f0d5d244703
[10] Fagundes-Moreira R, Bezerra-Santos M, Lia R, et al. Eyeworms of wild birds and new record of Thelazia (Thelaziella) aquilina (Nematoda: Spirurida). International Journal for Parasitology: Parasites and Wildlife. 2024. https://
[11] García-Livia K, Santana-Hernández KM, Udiz-Rodríguez R, et al. Thelazia callipaeda (Spirurida, Thelaziidae): First record of an imported ocular parasite in dogs from the Canary Islands, Spain. Veterinary Parasitology: Regional Studies and Reports. 2025. https://www.semanticscholar.org/paper/e6c797e3c9d5aa7325e1c9e7005481220500b69a
[12] Sobotyk C, Dietrich J, Verocai GG, et al. Thelazia callipaeda Eyeworms in American Black Bear, Pennsylvania, USA, 2023. Emerging Infectious Diseases. 2024. https://www.semanticscholar.org/paper/bc9527addbef0f914dd9d734d14ab2ace714ff9c
[13] Filip-Hutsch K, Bezerra-Santos M, Ricci A, et al. Epidemiological, morphological, and molecular characterization of Thelazia spp. in European bison (Bison bonasus) in Poland. Scientific Reports. 2025. https://www.semanticscholar.org/paper/1e44f770e6f5e8852d45a4a46dd6c80340744517
[14] Cotuțiu V, Cazan C, Ionicǎ A, et al. Thelazia rhodesi and Thelazia skrjabini in wild European bison (Bison bonasus) and farmed American bison (Bison bison) from Romania, 2021-2023. Journal of Wildlife Diseases. 2025. https://www.semanticscholar.org/paper/c3f1665fa29f5e8ae7af1e538da958ac16574433
[15] Han Z, Zhong Y, Chen NW, et al. Nematicidal Effects and Cytotoxicity of Levamisole on Thelazia callipaeda. Animals. 2025. https://www.semanticscholar.org/paper/7a37d4bd7d90dfc3ee358ae171b33a1708943944
[16] Martins AL, Stevens L, Kieninger M, et al. The genome sequence of a nematode, Thelazia callipaeda Railliet & Henry, 1910 (Rhabditida: Thelaziidae). Wellcome Open Research. 2025. https://www.semanticscholar.org/paper/368f41f609fe1adbef90606bc9d25d317629ce09
[17] Wang X, Li C. Thelazia callipaeda infestation in an 8-month-old male infant in Hubei, China. Wiener Klinische Wochenschrift. 2025. https://www.semanticscholar.org/paper/55525cbda222dcd06bba0c16cca6a722723ea722
[18] Li L, Sun W, Li C, et al. Human Thelazia callipaeda Infection: A Case Report. International Journal of Infectious Diseases. 2025. https://www.semanticscholar.org/paper/96b54c546a20fb7aab2daed1e6aa828f4c694671
[19] Gu X, Wang E, Li N, et al. Live Thelazia Callipaeda in the Vitrectomized Eye after Intraocular Surgery: A Case Report. International Journal of Infectious Diseases. 2025. https://www.semanticscholar.org/paper/dccf167af1be6ad5cd2fb15e79e065073c29c0f2
[20] Wang M, Ni T, Yu W. Thelazia callipaeda in Vitreous Cavity after Cataract Surgery. Ophthalmology. 2025. https://www.semanticscholar.org/paper/17cd4f80e7ce36ee57d021e2b96216a92995da12