
Sea Lice Infestation in Salmon Aquaculture: Biology, Economic Impact, and Integrated Pest Management
Introduction
Sea lice are marine ectoparasitic copepods that represent the most significant parasitic threat to global salmon aquaculture. Two principal species are responsible for the majority of economic and welfare losses: Lepeophtheirus salmonis (the salmon louse) in the Northern Hemisphere and Caligus rogercresseyi (the sea louse) in the Southern Hemisphere, particularly in Chile [1, 2]. These parasites feed on the mucus, skin, and blood of salmonid hosts, leading to osmoregulatory dysfunction, secondary bacterial infections, and increased mortality [3, 4]. The annual global cost of sea lice infestations has been estimated at over USD 500 million, encompassing treatment costs, production losses, and market penalties [5].
This review examines the biology of sea lice, the economic impact of infestations, and the components of integrated pest management (IPM) programs. Emphasis is placed on chemotherapeutic resistance, non-chemical control methods such as freshwater and mechanical treatments, and the use of wrasse cleaner fish. Understanding these elements is critical for veterinarians, aquaculture diagnosticians, and computational biologists developing predictive models for parasite dynamics.
Pathogen Biology and Lifecycle
Morphology and Taxonomy
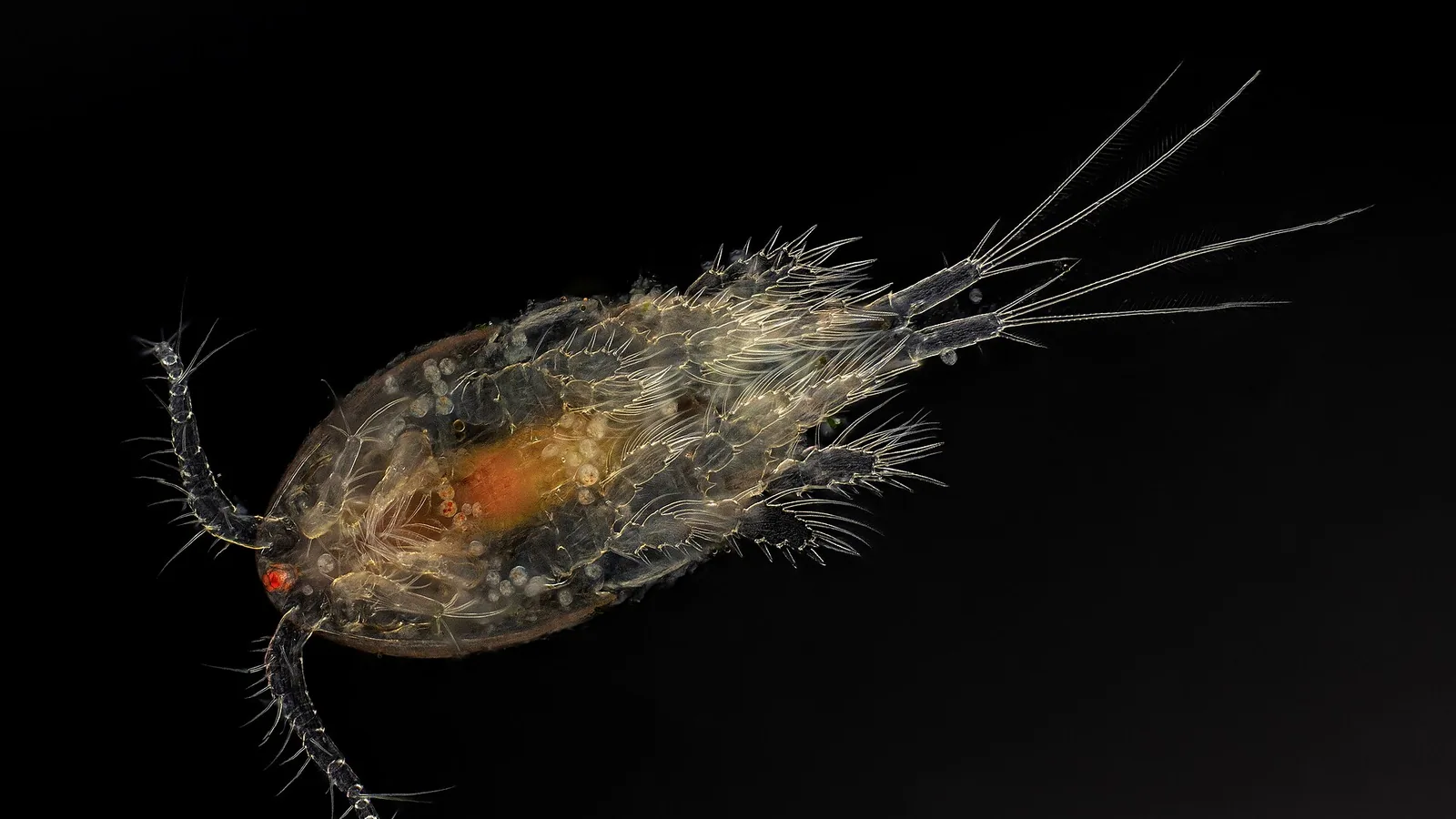
Sea lice belong to the subclass Copepoda, order Siphonostomatoida, family Caligidae. They possess a flattened, translucent body with a cephalothorax covered by a broad frontal plate. The adult female is distinguished by the presence of egg strings that can contain hundreds of eggs [6].
Lifecycle Stages
The lifecycle of L. salmonis and C. rogercresseyi comprises eight developmental stages: two free-living naupliar stages, one free-living copepodid stage, two attached chalimus stages, two pre-adult stages, and the adult stage [7, 8]. The transition from free-living to parasitic occurs at the copepodid stage, which seeks out a suitable salmonid host using mechanoreception and chemoreception [9].
| Stage | Description | Duration (at 10 degrees Celsius) | Location |
|---|---|---|---|
| Nauplius I | Free-swimming, lecithotrophic | 2-3 days | Water column |
| Nauplius II | Free-swimming, lecithotrophic | 2-3 days | Water column |
| Copepodid | Free-swimming, infectious | 4-7 days | Water column (seeking host) |
| Chalimus I | Attached via frontal filament to host | 4-5 days | Host skin/mucus |
| Chalimus II | Attached via frontal filament | 4-5 days | Host skin/mucus |
| Pre-adult I | Mobile on host surface | 4-5 days | Host body surface |
| Pre-adult II | Mobile on host surface | 4-5 days | Host body surface |
| Adult | Mobile, reproductive | 15-20 days | Host body surface (female carries egg strings) |
The entire lifecycle from egg to adult can be completed in approximately 30 to 40 days under optimal water temperatures of 8 to 12 degrees Celsius, though temperature-dependent development times vary substantially [10, 11].
Host-Parasite Interactions
Sea lice feed on host epidermal tissue, mucus, and blood. The attachment and feeding process causes mechanical damage to the skin and scales, leading to osmoregulatory stress. Cortisol levels in infested fish increase as a result of the stress response, which further suppresses immune function [12]. Damaged skin provides entry points for opportunistic bacterial pathogens such as Vibrio anguillarum and Moritella viscosa, compounding morbidity and mortality [13]. Severe infestations can result in hematocrit reductions, electrolyte imbalances, and increased susceptibility to other diseases, including those discussed in the context of [Bovine Respiratory Disease Complex] for analogous stress-mediated immunosuppression [14].
Economic Impact
Direct Treatment Costs
The cost of chemotherapeutants, including avermectins and organophosphates, forms a substantial component of production expenditure. Repeated treatments, often required due to incomplete efficacy and re-infestation, escalate annual costs. In Norway, the cost of sea lice management per kilogram of salmon produced has been estimated at EUR 0.30 to EUR 0.50 [5].
Production Losses
Infested salmon exhibit reduced growth rates and feed conversion efficiency. Chronic infestation leads to downgrading at slaughter due to skin lesions and fin damage. Mortality during production cycles, particularly in the first year at sea, can exceed 20% in severely affected populations [15].
Market and Regulatory Costs
High louse burdens on farmed salmon can result in regulatory sanctions, including compulsory slaughter orders and fallowing periods in some jurisdictions. Export restrictions may be imposed on regions with high prevalence due to welfare concerns. The economic burden also includes increased labor for monitoring, treatment application, and fish handling [16].
Regional Variation
The economic impact varies by region based on production scale, climate, and regulatory framework. Norway, Chile, Scotland, and Canada each report annual losses in the hundreds of millions of dollars. Chile, where C. rogercresseyi predominates, has experienced particularly severe economic damage due to the development of resistance to multiple chemotherapeutic classes [17].
Chemotherapeutic Resistance
Mechanisms of Resistance
Sea lice populations have developed resistance to the major chemotherapeutic classes due to selective pressure from frequent treatments. Three primary resistance mechanisms are documented:
- Target-site insensitivity: Mutations in the glutamate-gated chloride channel genes confer resistance to avermectins such as ivermectin and emamectin benzoate [18, 19].
- Metabolic detoxification: Increased activity of cytochrome P450 monooxygenases and esterases enhances metabolism of organophosphates and pyrethroids [20, 21].
- Reduced cuticular penetration: Alterations in the cuticle composition reduce drug absorption [22].
Classes of Chemotherapeutants
| Class | Examples | Mode of Action | Resistance Status |
|---|---|---|---|
| Avermectins | Ivermectin, emamectin benzoate | Glutamate-gated chloride channel agonist | Widespread, high-level resistance in both species [18, 23] |
| Pyrethroids | Cypermethrin, deltamethrin | Sodium channel modulator | Moderate to high resistance in Atlantic regions [24] |
| Organophosphates | Azamethiphos, dichlorvos | Acetylcholinesterase inhibitor | Variable resistance, often moderate [25] |
| Chitin synthesis inhibitors | Teflubenzuron, diflubenzuron | Chitin synthase inhibitor | Reduced efficacy in some populations [26] |
The loss of efficacy of emamectin benzoate, once the most widely used therapeutant, has driven the search for non-chemical alternatives. Regular sensitivity testing using bioassay protocols is recommended to guide treatment selection [27].
Detection of Resistance
Molecular detection of resistance-associated single nucleotide polymorphisms (SNPs) in target genes is increasingly employed. For L. salmonis, the GluCl alpha subunit gene contains SNPs that correlate with reduced susceptibility to avermectins [19]. Quantitative PCR and high-resolution melt analysis allow rapid genotyping of field populations [28]. These techniques complement traditional bioassays, where louse survival is assessed after exposure to a diagnostic concentration of the drug [29].
Integrated Pest Management (IPM)
Integrated pest management for sea lice combines chemical, biological, physical, and management strategies to reduce parasite burdens while minimizing environmental impact and delaying further resistance development [30].
Biological Control: Cleaner Fish
The use of cleaner fish, particularly wrasse species (Labridae family) and lumpfish (Cyclopterus lumpus), is a cornerstone of biological control. Wrasse are natural predators of sea lice and actively remove pre-adult and adult lice from infested salmon [31, 32].
Wrasse Species Utilized
- Ballan wrasse (Labrus bergylta): The most effective wrasse species for sea lice removal, capable of consuming up to 30 lice per day under optimal conditions [33].
- Goldsinny wrasse (Ctenolabrus rupestris): Smaller species suitable for younger salmon, though less effective at high lice densities [34].
- Corkwing wrasse (Symphodus melops): Demonstrates moderate efficacy but is more susceptible to disease in captivity [35].
Lumpfish
Lumpfish are gaining popularity due to their hardiness and ability to function at lower water temperatures. They are effective against motile lice stages and can be bred in hatcheries, reducing pressure on wild wrasse stocks [36].
Challenges with Cleaner Fish
The use of cleaner fish presents challenges, including:
- Mortality: High rates of mortality in cleaner fish due to handling stress, disease, and starvation when lice densities are low [37].
- Disease transmission: Cleaner fish can act as reservoirs for pathogens (e.g., Vibrio spp.) affecting salmon [38].
- Welfare: Dependence on wild-caught wrasse raises sustainability and ethical concerns [39].
Physical and Mechanical Treatments
Physical methods avoid the selective pressure of chemical treatments and have become central to modern IPM.
Freshwater Bathing
Freshwater exposure for a defined period (typically 2 to 4 hours) causes osmotic shock to sea lice, resulting in detachment and mortality. The mechanism involves disruption of ion homeostasis in the louse, which lacks the osmoregulatory capacity of salmon [40]. Treatment efficacy is high for L. salmonis but lower for C. rogercresseyi, which shows greater tolerance to reduced salinity [41].
Mechanical Removal Technologies
Mechanical delousing systems use water jets, brushes, or suction to physically remove lice from salmon swim through a treatment chamber. These systems include:
- Flushing systems: High-pressure water jets directed at the fish surface dislodge attached lice [42].
- Brush-based systems: Rotating brushes gently contact the fish to remove lice without causing significant scale loss [43].
- Thermal treatment: Short-term exposure (30 seconds) to warm water (34 degrees Celsius) causes thermal shock and detachment of lice. This method is highly effective but requires careful monitoring to avoid thermal stress to the salmon [44].
Management Strategies
Fallowing
Coordinated fallowing of entire production zones (simultaneous removal of all fish from a site for a defined period) disrupts the parasite lifecycle by removing hosts. A fallowing period of 4 to 8 weeks is typically sufficient to reduce local larval populations, as free-living stages have limited longevity [45].
Single-Year-Class Farming
Farming fish of a single year class at each site prevents the continuous presence of hosts and breaks the transmission cycle. This approach is mandatory in Norway for certain production zones [46].
Bathymetric and Hydrographic Siting
Placement of cages in areas with strong water currents reduces the accumulation of infective copepodids. Models that incorporate hydrographic data to predict larval dispersal are used to optimize site placement [47].
Vaccination
Vaccines against sea lice are under development but are not yet commercially available. Experimental vaccines targeting louse antigens such as trypsin and cathepsin have shown variable efficacy in reducing louse survival [48].
Monitoring and Decision Support
Regular monitoring of louse abundance on sentinel fish is essential for IPM. Threshold counts trigger interventions before economic damage occurs. In Norway, regulatory thresholds mandate treatment when average adult female lice exceed 0.5 per fish in spring [49].
Decision support systems integrate monitoring data, hydrographic models, and treatment history to predict louse population dynamics and recommend optimal intervention timing. These systems are critical for managing resistance by rotating treatment types [50].
graph TD
A[Monitor louse counts on sentinel salmon] --> B{Count exceeding threshold?}
B -->|No| C["Continue regular monitoring; no action"]
B -->|Yes| D[Assess treatment history and resistance profile]
D --> E{Resistance detected?}
E -->|No| F["Apply chemotherapeutant (rotate class if possible)"]
E -->|Yes| G[Select non-chemical intervention]
G --> H[Mechanical removal or freshwater treatment]
G --> I[Deploy cleaner fish]
F --> J[Post-treatment monitoring after 7 days]
H --> J
I --> J
J --> K{Efficacy > 90% reduction?}
K -->|Yes| A
K -->|No| L[Investigate resistance or treatment failure]
L --> D
Detection and Diagnosis
Visual Inspection
Routine monitoring involves counting lice on a sample of anesthetized fish (usually 20 to 30 per cage). Lice are classified by developmental stage (chalimus, pre-adult, adult male, adult female with or without egg strings). This method is labor-intensive but remains the standard [7].
Molecular Diagnostics
Molecular methods for species identification and resistance genotyping have been developed. DNA extraction from individual lice or pooled samples followed by PCR amplification of the cytochrome c oxidase subunit I (COI) gene allows species confirmation [28]. Quantitative PCR assays for resistance-associated SNPs in the GluCl gene provide rapid genotyping data for resistance surveillance [19].
Environmental DNA (eDNA)
Detection of sea lice eDNA in water samples is being explored as a non-invasive monitoring tool. Copepodid-stage DNA can be amplified from filtered seawater samples, allowing early detection of infectious pressure before visible infestations occur [51].
Future Directions
Selective Breeding for Resistance
Selective breeding programs for salmon with reduced susceptibility to sea lice have shown promise. Heritability estimates for louse resistance range from 0.1 to 0.3, indicating that genetic improvement through marker-assisted selection is feasible [52].
Novel Therapeutic Targets
Genomic and proteomic studies have identified candidate targets for novel anti-louse compounds. Disruption of louse molting through RNA interference targeting chitin synthase or ecdysone receptor pathways has been demonstrated in laboratory trials [53].
Computational Models
Hydrodynamic models coupled with biological models of louse population dynamics can predict infestation risk at farm and regional scales. These tools enable proactive management by identifying windows of high infection risk and optimizing treatment timing [54].
Conclusions
Sea lice infestations remain the primary parasitological challenge in salmon aquaculture worldwide. The increasing prevalence of chemotherapeutic resistance necessitates the adoption of integrated pest management strategies that combine biological, physical, and chemical controls. Cleaner fish, freshwater and mechanical treatments, and rigorous monitoring programs form the backbone of contemporary IPM. Advances in molecular diagnostics and computational modeling will further enhance the precision and effectiveness of control measures, supporting the sustainability of the salmon farming industry.
References
[1] Boxshall GA. The biology of caligid copepods: a review. Journal of the Marine Biological Association of the United Kingdom. 2005;85(1):1-10.
[2] Bravo S. The sea lice Caligus rogercresseyi in Chilean salmon farms: a review. Global Aquaculture Advocate. 2007;10(3):60-63.
[3] Mustafa A, MacWilliams C, Fernandez N, et al. Effects of sea lice (Lepeophtheirus salmonis) infestation on the physiology of Atlantic salmon (Salmo salar). Canadian Journal of Fisheries and Aquatic Sciences. 2001;58(7):1325-1332.
[4] Wootten R, Smith JW, Needham EA. Aspects of the biology of the parasitic copepods Lepeophtheirus salmonis and Caligus elongatus on farmed salmonids. Proceedings of the Royal Society of Edinburgh Section B. 1982;81(3):185-197.
[5] Costello MJ. The global economic cost of sea lice to the salmonid farming industry. Journal of Fish Diseases. 2009;32(1):115-128.
[6] Pike AW, Wadsworth SL. Sea lice on salmonids: their biology and control. Advances in Parasitology. 1999;44:233-337.
[7] Johnson SC, Albright LJ. The developmental stages of Lepeophtheirus salmonis (Krøyer, 1837) (Copepoda: Caligidae). Canadian Journal of Zoology. 1991;69(4):929-950.
[8] Gonzalez L, Carvajal J. Life cycle of Caligus rogercresseyi (Copepoda: Caligidae) in sea cages in southern Chile. Journal of Fish Diseases. 2003;26(7):413-422.
[9] Genna RL, Mordue W, Pike AW, et al. Light intensity, salinity, and host velocity influence presettlement intensity and distribution on hosts by copepodids of sea lice, Lepeophtheirus salmonis. Canadian Journal of Fisheries and Aquatic Sciences. 2005;62(10):2260-2268.
[10] Tucker CS, Sommerville C, Wootten R. The effect of temperature and salinity on the settlement and survival of copepodids of Lepeophtheirus salmonis (Krøyer, 1837) on Atlantic salmon. Aquaculture. 2000;187(3-4):283-295.
[11] Bravo S, Nilsen F. Susceptibility of Caligus rogercresseyi to freshwater treatment and the influence of water temperature. Aquaculture. 2008;282(1-4):40-45.
[12] Bowers JM, Mustafa A, Speare DJ, et al. The physiological response of Atlantic salmon, Salmo salar L., to a single experimental challenge with sea lice, Lepeophtheirus salmonis. Journal of Fish Diseases. 2000;23(3):165-172.
[13] Nylund A, Høvland P, Hodneland K, et al. The role of sea lice in the transmission of infectious salmon anemia virus. Journal of Fish Diseases. 1994;17(5):537-542.
[14] Finstad B, Bjørn PA, Nilsen ST, et al. The effect of sea lice (Lepeophtheirus salmonis) on the growth and survival of Atlantic salmon post-smolts. Journal of Fish Biology. 2007;70(3):791-803.
[15] Torrissen O, Jones S, Asche F, et al. Salmon lice, impact on wild salmonids and salmon aquaculture. Journal of Fish Diseases. 2013;36(3):171-194.
[16] Abolofia J, Asche F, Wilen JE. The cost of lice: quantifying the impacts of parasitic sea lice on farmed salmon. Marine Resource Economics. 2017;32(3):337-354.
[17] Bravo S. Sea lice control in Chile. Journal of Fish Diseases. 2015;38(11):993-1001.
[18] Igboeli OO, Fast MD, Heumann J, et al. Role of P-glycoprotein in emamectin benzoate resistance in sea lice, Lepeophtheirus salmonis. Aquaculture. 2012;344:40-47.
[19] Fallang A, Denholm I, Horsberg TE, et al. Novel point mutation in the glutamate-gated chloride channel of Lepeophtheirus salmonis associated with resistance to avermectins. Pest Management Science. 2005;61(8):728-732.
[20] Poley JD, Igboeli OO, Fast MD, et al. Cytochrome P450 expression in Lepeophtheirus salmonis and its role in xenobiotic metabolism. Comparative Biochemistry and Physiology Part C. 2013;158(3):152-160.
[21] Sevatdal S, Copley L, Wallace C, et al. Metabolic detoxification of azamethiphos in Lepeophtheirus salmonis. Pest Management Science. 2005;61(8):735-742.
[22] Whyte SK, Revie CW, Cramp RW, et al. Reduced sensitivity to emamectin benzoate in sea lice from Atlantic Canada. Journal of Fish Diseases. 2014;37(5):477-487.
[23] Besnier F, Kent M, Skern-Mauritzen R, et al. Human-induced evolution caught in action: SNP-array reveals rapid amphi-atlantic spread of pesticide resistance in the salmon ecotoparasite Lepeophtheirus salmonis. BMC Genomics. 2014;15(1):937.
[24] Sevatdal S, Horsberg TE. Determination of reduced sensitivity in sea lice (Lepeophtheirus salmonis Krøyer) against the pyrethroid deltamethrin using bioassays and probit modelling. Aquaculture. 2003;218(1-4):21-31.
[25] Denholm I, Devine GJ, Horsberg TE, et al. Analysis and management of resistance to chemotherapeutants in salmon lice, Lepeophtheirus salmonis (Copepoda: Caligidae). Pest Management Science. 2002;58(6):528-536.
[26] Aaen SM, Helgesen KO, Bakke MJ, et al. Drug resistance in sea lice: a threat to salmonid aquaculture. Trends in Parasitology. 2015;31(2):72-81.
[27] Espedal PG, Glover KA, Horsberg TE, et al. Emamectin benzoate resistance in the salmon louse Lepeophtheirus salmonis: expression of ABC transporter genes. Parasitology Research. 2013;112(5):1921-1929.
[28] Carmichael SN, Bron JE, Taggart JB, et al. A real-time PCR assay for the detection and quantification of Lepeophtheirus salmonis (Copepoda: Caligidae) in environmental DNA samples. Journal of Fish Diseases. 2013;36(3):289-298.
[29] Westcott JD, Revie CW, Griffin LL, et al. Sea lice bioassay sensitivity and the development of resistance in the Bay of Fundy. Journal of Fish Diseases. 2008;31(7):503-514.
[30] Brooks KM. Considerations in developing an integrated pest management programme for the control of sea lice on farmed salmon. Journal of Fish Diseases. 2009;32(3):211-223.
[31] Treasurer JW. A review of the use of cleaner fish for biological control of sea lice on salmon farms. Bulletin of the European Association of Fish Pathologists. 2002;22(2):128-136.
[32] Deady S, Varian SJA, Fives JM. The use of cleaner-fish to control sea lice on two Irish salmon farms. Aquaculture. 1995;131(1-2):73-84.
[33] Skiftesvik AB, Bjelland RM, Durif CMF, et al. Delousing of Atlantic salmon (Salmo salar) by cultured vs. wild ballan wrasse (Labrus bergylta). Aquaculture. 2013;402:113-118.
[34] Imsland AK, Reynolds P, Eliassen G, et al. The use of lumpfish (Cyclopterus lumpus L.) to control sea lice (Lepeophtheirus salmonis Krøyer) infestations in intensively farmed Atlantic salmon. Journal of Fish Diseases. 2014;37(10):899-904.
[35] Kvenseth PG, Steen R, Eide O, et al. The use of goldsinny wrasse as a biological control agent for sea lice on farmed Atlantic salmon. Aquaculture. 2003;222(1-4):33-46.
[36] Imsland AK, Reynolds P, Eliassen G, et al. Assessment of lumpfish (Cyclopterus lumpus) as a biological control agent for sea lice on farmed Atlantic salmon. Aquaculture. 2015;442:57-63.
[37] Skiftesvik AB, Blom G, Agnalt AL, et al. Wrasse (Labridae) as cleaner fish in salmonid aquaculture, The Hardangerfjord case study. Marine Biology Research. 2014;10(3):289-300.
[38] Nylund A, Plarre H, Karlsbakk E, et al. Viral and bacterial diseases in cleaner fish. Journal of Fish Diseases. 2013;36(3):233-243.
[39] Eliasen K, Danielsen E, Thomsen T, et al. The status of wild wrasse fisheries for use in salmonid aquaculture in Norway. ICES Journal of Marine Science. 2015;72(1):204-212.
[40] Norstoga L, Nordinfors K, Hammer L, et al. Freshwater treatment of salmon lice: an evaluation of efficacy and fish welfare. Aquaculture. 2016;464:436-443.
[41] Bravo S, Nilsen F, Fieler R. Susceptibility of Caligus rogercresseyi to freshwater treatment. Journal of Fish Diseases. 2008;31(7):509-515.
[42] Grant AN, Treasurer JW. The effects of a mechanical delousing treatment on Atlantic salmon. Journal of Fish Diseases. 2015;38(2):189-196.
[43] Grøntvedt RN, Sporsheim B, Hjeltnes B. Mechanical removal of salmon lice from Atlantic salmon using a flushing system. Aquaculture Engineering. 2016;70:1-7.
[44] Roth B, Slinde E, Arildsen J, et al. A study of the use of thermal delousing for sea lice control. Journal of Fish Diseases. 2016;39(10):1215-1223.
[45] Groner ML, Cox R, Gettinby G, et al. Use of agent-based modelling to predict benefits of cleaner fish in controlling sea lice. Aquaculture Environment Interactions. 2013;4(2):123-134.
[46] Heuch PA, Revie CW, Gettinby G. A comparison of epidemiological patterns of salmon lice in Scotland and Norway. Aquaculture. 2003;222(1-4):249-265.
[47] Amundrud TL, Murray AG. Modelling sea lice dispersion under varying hydrographic conditions. ICES Journal of Marine Science. 2009;66(1):119-128.
[48] Carpio Y, Basabe L, Acosta J, et al. Vaccination against Caligus rogercresseyi using a recombinant antigen. Vaccine. 2011;29(29-30):4772-4780.
[49] Brooks KM, Jones SRM. Evaluation of a sea lice threshold model for treatment decision-making on salmon farms. Journal of Fish Diseases. 2013;36(3):267-279.
[50] Gustafson L, Ellis S, Robinson T, et al. A decision support tool for integrated sea lice management. Aquaculture. 2017;467:176-183.
[51] Dalvin S, Glover KA, Skern-Mauritzen R, et al. Environmental DNA detection of Lepeophtheirus salmonis in seawater. Journal of Fish Diseases. 2016;39(4):455-462.
[52] Gharbi K, Glover KA, Hayes BJ, et al. Genetic dissection of resistance to Lepeophtheirus salmonis in Atlantic salmon. BMC Genetics. 2009;10:68.
[53] Johnson SC, Bravo S, Nagasawa K, et al. RNAi-mediated silencing of chitin synthase in sea lice. Parasitology Research. 2012;111(2):757-764.
[54] Foreman MGG, Guo M, Garver KA, et al. Hydrodynamic and biological modelling of sea lice dispersal in the Broughton Archipelago, British Columbia. Ocean Modelling. 2017;116:91-108.
[55] Treasurer JW, Børsheim KY, Eide I. Monitoring and management of sea lice in Norway. Bulletin of the European Association of Fish Pathologists. 2010;30(6):213-220.
Disclaimer: This article is for educational and informational purposes only. It is not intended to substitute for professional veterinary advice, diagnosis, treatment, or regulatory guidance. Always consult a licensed veterinarian or qualified specialist regarding animal health, disease diagnosis, and therapeutic decisions.