
Sea Lice (Lepeophtheirus salmonis) in Salmon Aquaculture: Integrated Pest Management
Introduction
The salmon louse Lepeophtheirus salmonis (Krøyer, 1837) is a marine ectoparasitic copepod that represents the most economically damaging pathogen in global salmonid aquaculture. Infestations cause direct pathological damage to host fish, increase susceptibility to secondary infections, and impose severe regulatory and market constraints on production. The global cost of sea lice management, including treatment expenses and production losses, is estimated to exceed USD 500 million annually [1, 2]. This review provides a comprehensive examination of L. salmonis biology, diagnostic methodologies, chemoresistance mechanisms, and the integrated pest management (IPM) framework that combines biological, chemical, physical, and genetic control strategies.
Taxonomy and Morphology
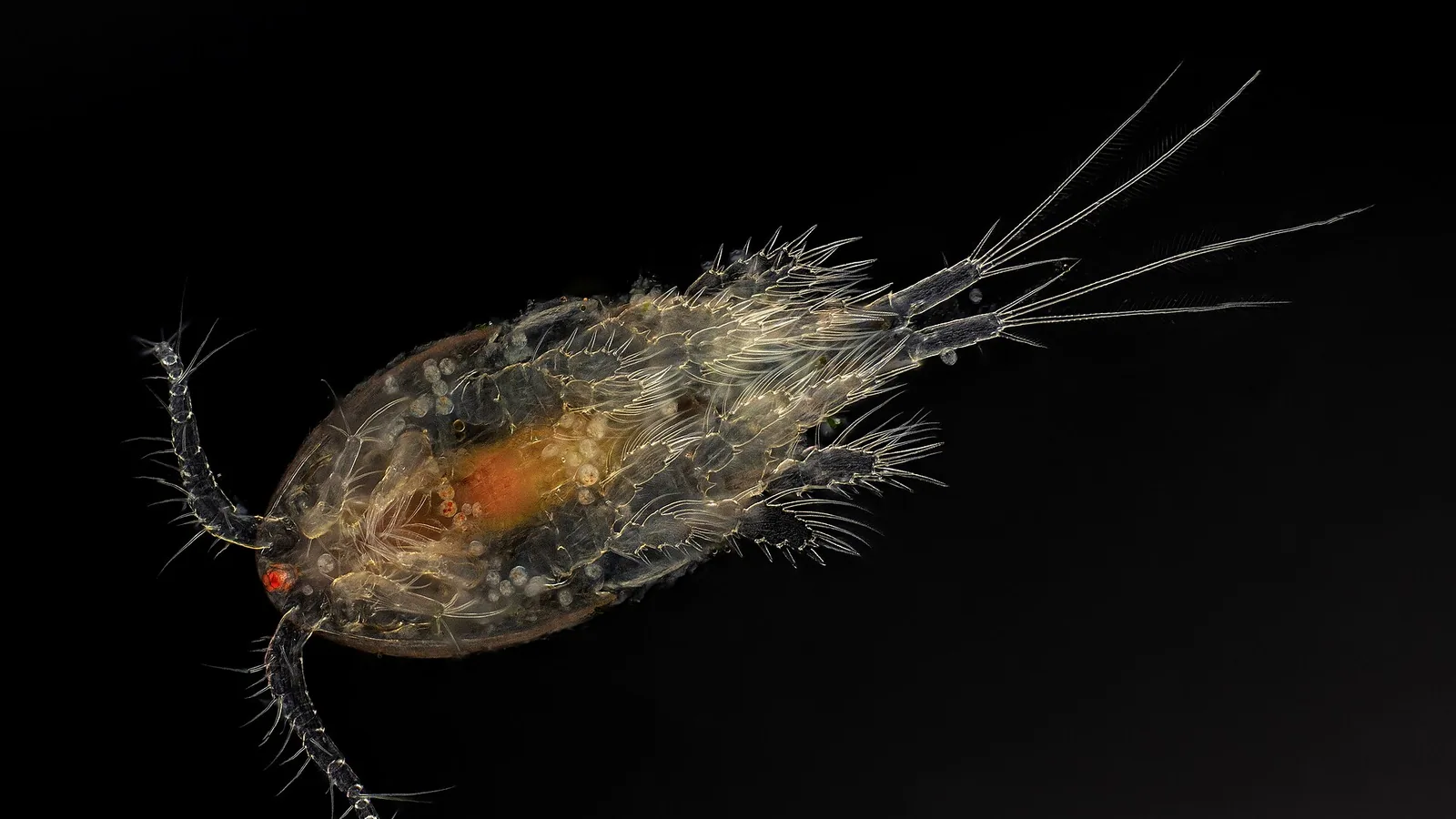
Lepeophtheirus salmonis belongs to the subclass Copepoda, order Siphonostomatoida, family Caligidae. The genus Lepeophtheirus contains over 100 species, but L. salmonis is the primary pathogen of Atlantic salmon (Salmo salar) and rainbow trout (Oncorhynchus mykiss) in the Northern Hemisphere [3]. A closely related species, Caligus rogercresseyi, dominates in the Southern Hemisphere (Chilean salmon aquaculture) and is often discussed in parallel literature.
The adult female L. salmonis measures 10 to 12 mm in total length, while males are smaller at 5 to 7 mm [4]. The body is dorsoventrally flattened with a cephalothorax that incorporates the first three thoracic segments. The frontal plates bear a lunule, a specialized adhesive structure unique to caligids. The fourth thoracic segment and genital complex are distinct, with the female possessing paired egg strings that can contain 500 to 1000 eggs per string [5].
Life Cycle
The L. salmonis life cycle comprises eight developmental stages separated by molts: two nauplius stages (I and II), one copepodid stage, four chalimus stages (I to IV), and two adult stages (preadult I and II, then adult) [6]. The life cycle is temperature dependent. At 10 degrees Celsius, the entire cycle from egg to adult requires approximately 40 days [7].
Free-Living Stages
Egg strings are extruded by adult females and remain attached until hatching. Nauplius I and II are nonfeeding, planktonic stages that rely on endogenous yolk reserves. The nauplius II molts into the copepodid stage, which is the infective stage. The copepodid is a free-swimming, phototactic, and rheotactic larva that must locate and attach to a suitable salmonid host within 4 to 7 days before energy reserves are depleted [8].
Parasitic Stages
Upon host attachment, the copepodid molts into the chalimus I stage. Chalimus stages I through IV are sessile, attached to the host by a frontal filament secreted from the frontal gland. These stages feed on host mucus, skin, and blood. The chalimus IV molts into the mobile preadult I stage, which is capable of moving freely across the host body surface. Preadult I and II stages and adults are highly mobile and feed aggressively [9].
Mating and Reproduction
Mating occurs on the host. The male grasps the female using his second antennae and transfers a spermatophore. Females can store sperm and produce multiple egg string pairs without remating. Fecundity is positively correlated with female body size and negatively correlated with water temperature [10].
Pathogenesis and Economic Impact
Direct Pathological Effects
Lepeophtheirus salmonis feeds on host mucus, epidermis, and blood. Attachment and feeding cause mechanical erosion of the epidermis, leading to osmoregulatory dysfunction, dermal ulceration, and stress. Heavy infestations (greater than 0.5 lice per gram of fish body weight) induce severe anemia, hypoproteinemia, and electrolyte imbalance [11]. In Atlantic salmon, chronic infestation suppresses the hypothalamic-pituitary-interrenal axis, elevating plasma cortisol levels and impairing immune function [12].
Secondary Infections
Lice-inflicted skin lesions serve as portals for secondary bacterial and viral infections. Coinfections with Aeromonas salmonicida (furunculosis), Vibrio anguillarum (vibriosis), and Moritella viscosa (winter ulcer disease) are well documented [13]. Viral pathogens such as infectious salmon anemia virus (ISAV) and salmonid alphavirus (SAV) show increased transmission rates in louse-infested populations [14]. The role of L. salmonis as a mechanical vector for ISAV has been demonstrated under experimental conditions [15].
Economic Losses
Economic losses arise from direct mortality, reduced growth rates, feed conversion inefficiency, treatment costs, and downgrading of fillet quality at processing. A meta-analysis of Norwegian salmon production estimated that each adult female louse on a farmed salmon reduces harvest weight by approximately 1.5 percent [16]. Regulatory thresholds in Norway, Scotland, Canada, and Chile mandate treatment when adult female counts exceed 0.1 to 0.5 lice per fish, depending on jurisdiction and season [17].
Diagnostic Methods
Visual Counting and Staging
The standard diagnostic method for sea lice infestation is manual counting and staging of lice on anaesthetized or sacrificed fish. Fish are netted, anaesthetized with tricaine methanesulfonate (MS-222), and examined under bright light. Lice are classified as copepodids, chalimus, preadults, or adults, and sex is recorded for adults [18]. This method is labor intensive, subject to observer bias, and underestimates early-stage infestations.
Molecular Diagnostics
Quantitative polymerase chain reaction (qPCR) assays targeting the internal transcribed spacer 1 (ITS-1) region of ribosomal DNA have been developed for species-specific detection of L. salmonis [19]. Environmental DNA (eDNA) approaches using qPCR can detect planktonic nauplii and copepodids in water samples, enabling early warning of infestation pressure [20]. Multiplex qPCR panels that simultaneously detect L. salmonis and C. rogercresseyi are available for regions where both species co-occur [21].
Hydrodynamic and Acoustic Sensors
Automated underwater camera systems coupled with machine learning algorithms can classify and count attached lice on free-swimming salmon in sea cages. These systems use convolutional neural networks trained on annotated image libraries and achieve sensitivity comparable to manual counting for adult stages [22]. Acoustic backscatter sensors have been deployed to estimate planktonic louse abundance in the water column [23].
Chemical Control and Resistance
Approved Chemotherapeutants
Chemical delousing agents are classified as bath treatments or in-feed treatments. Bath treatments include hydrogen peroxide (H2O2), azamethiphos (an organophosphate), and deltamethrin (a synthetic pyrethroid). In-feed treatments include emamectin benzoate (a macrocyclic lactone), teflubenzuron, and diflubenzuron (chitin synthesis inhibitors) [24].
Mechanisms of Resistance
Resistance to emamectin benzoate is the most widespread and economically significant. Resistance is associated with reduced target site sensitivity at glutamate-gated chloride channels and enhanced drug efflux via P-glycoprotein transporters [25]. Field studies in Norway, Scotland, and Canada have documented resistance ratios exceeding 100-fold compared to baseline susceptible populations [26].
Resistance to hydrogen peroxide involves upregulation of catalase and glutathione peroxidase enzymes that detoxify reactive oxygen species [27]. Pyrethroid resistance (deltamethrin) is linked to knockdown resistance (kdr) mutations in the voltage-gated sodium channel gene [28].
Resistance Monitoring
Bioassays using dose-response curves are the standard method for resistance detection. The maximum efficacy test (MET) exposes lice to a single discriminating concentration of a chemotherapeutant and records mortality after 24 to 48 hours [29]. Molecular assays targeting known resistance alleles, such as allele-specific PCR for the GluCl alpha subunit gene, are under development for rapid genotyping [30].
Biological Control: Cleaner Fish
Cleaner Fish Species
Biological control using cleaner fish is a cornerstone of IPM in Norwegian and Scottish salmon aquaculture. The primary species used are the lumpfish (Cyclopterus lumpus) and the ballan wrasse (Labrus bergylta). Other wrasse species including goldsinny wrasse (Ctenolabrus rupestris), corkwing wrasse (Symphodus melops), and rock cook (Centrolabrus exoletus) are also deployed [31].
Efficacy and Limitations
Cleaner fish actively remove preadult and adult L. salmonis from host salmon. Lumpfish are effective at water temperatures between 4 and 14 degrees Celsius, while ballan wrasse require temperatures above 8 degrees Celsius [32]. Efficacy is influenced by cleaner fish size, stocking density, and the availability of alternative feed. Mortality of cleaner fish in sea cages can exceed 50 percent due to handling stress, starvation, and disease [33].
Welfare and Disease Concerns
Cleaner fish are susceptible to bacterial pathogens including Atypical Aeromonas salmonicida and Vibrio spp., as well as parasitic infections such as Neoparamoeba perurans (amoebic gill disease) [34]. Vaccination protocols for cleaner fish are being developed but are not yet standardized. The welfare of cleaner fish is a regulatory concern in Norway, where guidelines mandate minimum size, feeding regimes, and health monitoring [35].
Physical and Mechanical Control
Thermal Delousing
Thermal delousing exposes fish to seawater heated to 28 to 34 degrees Celsius for 20 to 30 seconds. The thermal shock causes detachment of attached lice without causing significant mortality to the salmon [36]. Commercial thermal delousing vessels process up to 300 tons of fish per hour. Concerns regarding thermal stress and gill pathology in salmon have been raised, particularly at higher temperatures and during summer months [37].
Hydrolicer and Mechanical Delousing
The Hydrolicer system uses water jets and physical brushing to dislodge lice. Fish pass through a series of chambers where high-pressure water nozzles and soft rubber brushes remove attached lice. Efficacy ranges from 70 to 95 percent for adult lice, with lower efficacy for chalimus stages [38].
Freshwater and Low-Salinity Baths
Freshwater bathing for 2 to 4 hours causes osmotic shock to L. salmonis, which is a stenohaline marine organism. This method is used in freshwater hatcheries and during transfer of smolts to sea. Low-salinity brackish water (less than 15 parts per thousand) also reduces louse survival and fecundity [39].
Selective Breeding for Resistance
Heritability of Resistance
Quantitative genetic studies have demonstrated that resistance to L. salmonis infestation is moderately heritable in Atlantic salmon. Estimates of heritability (h2) for louse count range from 0.12 to 0.33 depending on population and statistical model [40]. Genomic selection using single nucleotide polymorphism (SNP) arrays can increase the accuracy of breeding values for louse resistance [41].
Quantitative Trait Loci
Genome-wide association studies (GWAS) have identified several quantitative trait loci (QTL) associated with louse resistance on chromosomes 3, 6, and 17 of Atlantic salmon [42]. Candidate genes in these regions include major histocompatibility complex (MHC) class I and class II genes, as well as genes involved in the interferon gamma pathway [43].
Implementation in Breeding Programs
Commercial breeding programs in Norway and Scotland now include louse resistance as a selection trait. Estimated genetic gain per generation is 10 to 15 percent reduction in louse count. Genomic selection reduces the generation interval and accelerates genetic progress compared to pedigree-based selection.
Integrated Pest Management Framework
IPM for sea lice combines multiple control modalities to reduce reliance on any single intervention and delay the emergence of resistance. The IPM framework includes the following components.
Monitoring and Thresholds
Regular monitoring of louse abundance on farmed and wild salmon is essential. Action thresholds for treatment vary by jurisdiction. In Norway, the maximum allowable limit is 0.2 adult female lice per fish during the spring and summer months. Sentinel cages containing untreated fish are used to monitor infestation pressure in some regions.
Coordinated Fallowing
Regional coordination of fallowing periods (leaving sites empty for 4 to 8 weeks) disrupts the louse life cycle by removing the host population. Synchronized fallowing across management areas has been shown to reduce overall louse abundance and the frequency of chemical treatments.
Bath and In-Feed Treatment Rotation
Rotating between chemotherapeutants with different modes of action reduces selection pressure for resistance. A typical rotation schedule alternates between emamectin benzoate (in-feed), hydrogen peroxide (bath), and deltamethrin (bath) over successive treatment cycles.
Biological and Physical Integration
Cleaner fish are deployed prophylactically at stocking densities of 5 to 15 percent of salmon biomass. Thermal and mechanical delousing are used as salvage treatments when louse counts exceed thresholds. Selective breeding for resistance provides a cumulative, long-term reduction in susceptibility.
Vaccination and Immunostimulants
No commercial vaccine against L. salmonis is currently available. Experimental vaccines targeting the louse midgut proteins (e.g., myosin, tropomyosin) have shown partial efficacy in laboratory trials. Oral immunostimulants containing beta-glucans and nucleotides have been reported to reduce louse attachment rates by 20 to 30 percent.
Mermaid Diagram: IPM Decision Workflow
flowchart TD
A[Regular Louse Monitoring] --> B{Count > Threshold?}
B -->|No| C[Continue Monitoring]
B -->|Yes| D[Assess Resistance Status]
D --> E{Resistance Detected?}
E -->|No| F["Chemical Treatment<br>(Rotate MOA)"]
E -->|Yes| G[Non-Chemical Intervention]
G --> H[Thermal Delousing]
G --> I[Mechanical Delousing]
G --> J[Freshwater Bath]
F --> K[Post-Treatment Count]
H --> K
I --> K
J --> K
K --> L{Count Below Threshold?}
L -->|Yes| C
L -->|No| M[Deploy Cleaner Fish]
M --> N[Evaluate Cleaner Fish Health]
N --> O[Consider Fallowing]
O --> P[Restock with Resistant Strain]
P --> A
Conclusion
Lepeophtheirus salmonis remains the most significant parasitic threat to salmon aquaculture globally. The evolution of resistance to multiple chemotherapeutant classes has driven the development of an integrated pest management approach that combines biological control using cleaner fish, physical and thermal delousing, selective breeding for host resistance, and coordinated regional management. Advances in molecular diagnostics, including qPCR-based eDNA detection and SNP-based genomic selection, provide powerful tools for surveillance and long-term control. Continued research into vaccine development, cleaner fish welfare, and resistance allele monitoring will be essential to sustain the economic viability and environmental sustainability of salmon aquaculture.
References
[1] Costello MJ. The global economic cost of sea lice to the salmonid farming industry. Journal of Fish Diseases. 2009;32(1):115-118.
[2] Johnson SC, Treasurer JW, Bravo S, Nagasawa K, Kabata Z. A review of the impact of parasitic copepods on marine aquaculture. Zoological Studies. 2004;43(2):229-243.
[3] Boxshall GA, Defaye D. Global diversity of copepods (Crustacea: Copepoda) in freshwater. Hydrobiologia. 2008;595(1):195-207.
[4] Schram TA. Supplementary descriptions of the developmental stages of Lepeophtheirus salmonis (Krøyer, 1837) (Copepoda: Caligidae). Sarsia. 1993;78(3-4):197-209.
[5] Heuch PA, Nordhagen JR, Schram TA. Egg production in the salmon louse Lepeophtheirus salmonis (Krøyer) in relation to origin and water temperature. Aquaculture Research. 2000;31(10):805-814.
[6] Hamre LA, Eichner C, Caipang CMA, Dalvin ST, Bron JE, Nilsen F, et al. The salmon louse Lepeophtheirus salmonis (Copepoda: Caligidae) life cycle has only two chalimus stages. PLoS ONE. 2013;8(9):e73539.
[7] Samsing F, Oppedal F, Dalvin S, Johnsen I, Vågseth T, Dempster T. Salmon lice (Lepeophtheirus salmonis) development times, body size, and reproductive outputs follow universal temperature rules. Journal of Fish Biology. 2016;88(5):1841-1853.
[8] Heuch PA, Karlsen HE. Detection of infrasonic water oscillations by copepodids of Lepeophtheirus salmonis (Copepoda: Caligida). Journal of Plankton Research. 1997;19(6):735-747.
[9] Pike AW, Wadsworth SL. Sealice on salmonids: their biology and control. Advances in Parasitology. 1999;44:233-337.
[10] Nordhagen JR, Heuch PA, Schram TA. Size as indicator of origin of salmon lice Lepeophtheirus salmonis (Copepoda: Caligidae). Contributions to Zoology. 2000;69(1-2):99-108.
[11] Grimnes A, Jakobsen PJ. The physiological effects of salmon lice infection on post-smolt of Atlantic salmon. Journal of Fish Biology. 1996;48(6):1179-1194.
[12] Bowers JM, Mustafa A, Speare DJ, Conboy GA, Brimacombe M, Sims DE, et al. The physiological response of Atlantic salmon, Salmo salar L., to a single experimental challenge with sea lice, Lepeophtheirus salmonis. Journal of Fish Diseases. 2000;23(3):165-172.
[13] Mustafa A, MacWilliams C, Fernandez N, Matchett K, Conboy GA, Burka JF. Effects of sea lice (Lepeophtheirus salmonis Krøyer, 1837) infestation on macrophage functions in Atlantic salmon (Salmo salar L.). Fish and Shellfish Immunology. 2000;10(1):47-59.
[14] Barker SE, Bricknell IR, Covello J, Purcell S, Fast MD, Wolters W, et al. Sea lice, Lepeophtheirus salmonis, as a vector for infectious salmon anaemia virus (ISAV) in Atlantic salmon, Salmo salar L. Journal of Fish Diseases. 2019;42(5):731-740.
[15] Oelckers K, Vike S, Duesund H, Gonzalez J, Wadsworth S, Nylund A. The role of Lepeophtheirus salmonis in the transmission of infectious salmon anaemia virus. Journal of Fish Diseases. 2014;37(8):707-717.
[16] Abolofia J, Asche F, Wilen JE. The cost of lice: quantifying the impacts of parasitic sea lice on farmed salmon. Marine Resource Economics. 2017;32(3):329-349.
[17] Heuch PA, Bjørn PA, Finstad B, Holst JC, Asplin L, Nilsen F. A review of the Norwegian 'National Action Plan Against Salmon Lice on Salmonids': the effect on wild salmonids. Aquaculture. 2005;246(1-4):79-92.
[18] Treasurer JW, Pope JA. Selection of host sample number and design of a monitoring programme for ectoparasitic sea lice (Copepoda: Caligidae) on farmed Atlantic salmon, Salmo salar. Aquaculture. 2000;187(1-2):45-60.
[19] McBeath AJA, Penston MJ, Snow M, Cook PF, Bricknell IR, Cunningham CO. Development and application of real-time PCR for specific detection of Lepeophtheirus salmonis and Caligus elongatus larvae in Scottish plankton samples. Diseases of Aquatic Organisms. 2006;73(2):141-150.
[20] Dalvin S, Glover KA, Sørvik AGE, Seljestad GW, Taggart JB, Skugor S, et al. Environmental DNA monitoring of the salmon louse (Lepeophtheirus salmonis) in Norwegian fjords. ICES Journal of Marine Science. 2020;77(7-8):2835-2845.
[21] Núñez-Acuña G, Gallardo-Escárate C. Multiplex PCR for the simultaneous detection of the salmon louse Caligus rogercresseyi and Lepeophtheirus salmonis. Journal of Fish Diseases. 2013;36(11):947-955.
[22] Bui S, Oppedal F, Samsing F, Dempster T. Automated sea lice counting using deep learning. Aquacultural Engineering. 2020;89:102058.
[23] Patanasatienkul T, Sanchez J, Rees E, Pfeiffer DU, Revie CW. Acoustic detection of planktonic sea lice: a field evaluation. Aquaculture Environment Interactions. 2015;7(1):1-12.
[24] Burka JF, Hammell KL, Horsberg TE, Johnson GR, Rainnie DJ, Speare DJ. Drugs in salmonid aquaculture: a review. Journal of Veterinary Pharmacology and Therapeutics. 1997;20(5):333-349.
[25] Igboeli OO, Fast MD, Heumann J, Burka JF. Role of P-glycoprotein in emamectin benzoate (SLICE) resistance in sea lice, Lepeophtheirus salmonis. Aquaculture. 2012;344-349:40-47.
[26] Aaen SM, Helgesen KO, Bakke MJ, Kaur K, Horsberg TE. Drug resistance in sea lice: a threat to salmonid aquaculture. Trends in Parasitology. 2015;31(2):72-81.
[27] Helgesen KO, Horsberg TE. Single-dose field bioassay for sensitivity testing of sea lice to hydrogen peroxide. Journal of Fish Diseases. 2013;36(3):267-276.
[28] Kaur K, Helgesen KO, Bakke MJ, Horsberg TE. Mechanism behind resistance against the organophosphate azamethiphos in salmon lice (Lepeophtheirus salmonis). PLoS ONE. 2015;10(4):e0124220.
[29] Sevatdal S, Horsberg TE. Determination of reduced sensitivity in sea lice (Lepeophtheirus salmonis Krøyer) against the pyrethroid deltamethrin using bioassays and probit modelling. Aquaculture. 2003;218(1-4):21-31.
[30] Carmichael SN, Bron JE, Taggart JB, Ireland JH, Bekaert M, Burgess STG, et al. Identification of a single nucleotide polymorphism associated with emamectin benzoate resistance in the salmon louse Lepeophtheirus salmonis. International Journal for Parasitology: Drugs and Drug Resistance. 2013;3:60-68.
[31] Imsland AK, Reynolds P, Eliassen G, Hangstad TA, Foss A, Vikingstad E, et al. The use of lumpfish (Cyclopterus lumpus L.) to control sea lice (Lepeophtheirus salmonis Krøyer) infestations in intensively farmed Atlantic salmon (Salmo salar L.). Aquaculture. 2014;424-425:18-23.
[32] Skiftesvik AB, Bjelland RM, Durif CMF, Johansen IS, Browman HI. Delousing of Atlantic salmon (Salmo salar) by cultured vs. wild ballan wrasse (Labrus bergylta). Aquaculture. 2013;402-403:113-118.
[33] Treasurer JW. Mortality of farmed wrasse (Labridae) due to anaesthetics. Veterinary Record. 2002;151(19):570-571.
[34] Erkinharju T, Dalmo RA, Hansen M, Seternes T. Cleaner fish in aquaculture: review on diseases and vaccination. Fish and Shellfish Immunology. 2021;119:128-140.
[35] Norwegian Veterinary Institute. Health and welfare of cleaner fish used in Norwegian salmon aquaculture. Report Series. 2020;7:1-45.
[36] Grøntvedt RN, Nerbøvik IKG, Viljugrein H, Lillehaug A, Nilsen H, Gjevre AG. Thermal de-licing of salmonid fish: documentation of fish welfare. Aquaculture. 2015;441:1-8.
[37] Overton K, Dempster T, Oppedal F, Kristiansen TS, Gismervik K, Stien LH. Salmon lice treatments and salmon mortality in Norwegian aquaculture: a review. Reviews in Aquaculture. 2019;11(4):1038-1054.
[38] Stien LH, Lind MB, Oppedal F, Wright DW, Seternes T. Hydrolicer: a novel mechanical delousing method for Atlantic salmon. Aquacultural Engineering. 2018;83:1-8.
[39] Bricknell IR, Dalesman SJ, O'Shea B, Pert CC, Luntz AJ. Effect of environmental salinity on sea lice Lepeophtheirus salmonis settlement success. Diseases of Aquatic Organisms. 2006;71(3):201-212.
[40] Gjerde B, Ødegård J, Thorland I. Estimates of genetic variation in the susceptibility of Atlantic salmon (Salmo salar) to the salmon louse Lepeophtheirus salmonis. Aquaculture. 2011;314(1-4):66-72.
[41] Tsai HY, Hamilton A, Tinch AE, Guy DR, Bron JE, Taggart JB, et al. Genomic prediction of host resistance to sea lice in farmed Atlantic salmon populations. Genetics Selection Evolution. 2016;48(1):47.
[42] Gharbi K, Glover KA, Manahan LC, Bron JE, Taggart JB, Bekaert M, et al. The genetics of resistance to the salmon louse Lepeophtheirus salmonis in Atlantic salmon. BMC Genomics. 2015;16(1):1007.
[43] Robledo D, Gutierrez AP, Barría A, Lhorente JP, Houston RD, Yáñez JM. Discovery and functional annotation of quantitative trait loci affecting resistance to sea lice in Atlantic salmon. Frontiers in Genetics. 2019;10:56.
Disclaimer: This article is for educational and informational purposes only. It is not intended to substitute for professional veterinary advice, diagnosis, treatment, or regulatory guidance. Always consult a licensed veterinarian or qualified specialist regarding animal health, disease diagnosis, and therapeutic decisions.